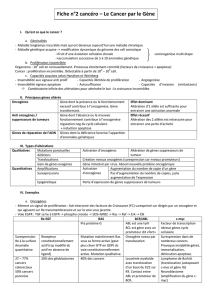
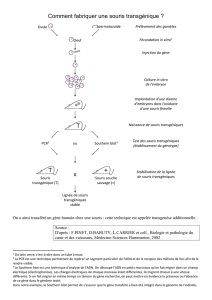
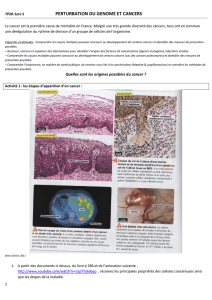
Les effets in vivo pas comparables de certains - iPubli

NOUVELLES
1992 ; 8 :
Les effets
pas comparables
à
à
à
à
de certains produits géniques ne sont
à ceux qu'on leur connaÎt
à
à
à
à
<<
1
à
à
à
Oe
s n ° 10 vol. 8, démb 92

à
à
à
à
à
à
à
s n ° 10 vol. 8, re 92
à
.
,
1
/
2
100%