revue générale Différentes approches de vaccination thérapeutique

revue générale
Différentes approches de vaccination
thérapeutique dans le traitement
de l’infection par le VIH-1
M. Lazrek
P.E. Lobert
A. Goffard
C. Schanen
A. Dewilde
L. Bocket
D. Hober
Service de virologie,
UPRES EA 3610, CHRU de Lille
Article reçu le 10 mai 2005,
accepté le 11 août 2005
Résumé.L’infection par le VIH-1 est une pandémie majeure. L’introduction
de multithérapies antirétrovirales a permis de réduire considérablement les
taux de morbidité et de mortalité liés au virus. Ces traitements ne permettent
néanmoins pas l’éradication du virus. Leur efficacité est notamment limitée par
leur complexité, leur toxicité à long terme et l’apparition de mutations de
résistance. La vaccination thérapeutique constitue une approche additionnelle
pour le traitement de l’infection par le VIH-1. Elle vise à amplifier et renforcer
les réponses immunitaires spécifiques anti-VIH. Différents immunogènes ont
été testés dans divers schémas thérapeutiques. Cet article propose une revue
des différentes stratégies adoptées pour la vaccination thérapeutique dans
l’infection par le VIH-1.
Mots clés :VIH, immunothérapie, vaccination
Abstract.HIV-1 infection is a major pandemic situation. With the advent of
highly active antiretroviral therapy (HAART), morbidity and mortality associa-
ted with HIV-1 infection have been dramatically reduced. However, HAART
does not enable eradication of the virus. The efficacy of these new regimens is
limited by problems over long-term use such as toxicity and resistance. Thera-
peutic vaccination is an alternative approach to HIV-1 infection. The main aim
is to boost and reinforce virus-specific host immune responses. Several immu-
nogens and schedules of immunization have been tested. In this review, various
strategies designed for therapeutic vaccines for HIV-1 infection are presented.
Key words:HIV, immunotherapy, vaccination
L’infection par le virus de l’immunodéficience humaine
(VIH) est une pandémie majeure. En 2004, l’Organisation
mondiale de la santé a estimé que près de 40 millions de
personnes étaient contaminées à travers le monde ; 5 mil-
lions étaient nouvellement infectées et 3 millions décédées
suite à la maladie [1].
La multiplication continue du VIH dans les lymphocytes T
CD4+ induit un déficit immunitaire progressif. La maladie
aboutit inexorablement chez la majorité des patients au
syndrome d’immunodéficience acquise (sida) et au décès
[2]. Un faible pourcentage de sujets infectés peut néan-
moins maintenir un contrôle immunitaire efficace du virus
pendant plus de 20 ans. Ces sujets sont qualifiés d’asymp-
tomatiques à long terme [3].
L’introduction en 1996 des multithérapies antirétrovirales
actives (HAART) a été responsable d’un bouleversement
du pronostic de l’infection. Ces traitements ont considéra-
blement réduit la morbidité et la mortalité liées au VIH. Ils
ne permettent néanmoins pas l’éradication du virus ni le
rétablissement d’un fonctionnement complètement normal
du système immunitaire [4]. Leur maintien à vie semble
donc nécessaire pour contrôler la réplication virale. Toute-
fois, leur utilisation à long terme se heurte à de nombreux
problèmes, liés notamment à leur toxicité, à l’apparition
de mutations de résistance et aux problèmes d’observance.
De plus, en raison de leur coût et de la complexité de leur
surveillance, ces médicaments restent inaccessibles à la
grande majorité des patients [5]. Ainsi, moins de 2 % des
patients infectés par le VIH à travers le monde ont accès
aux traitements anti-rétroviraux [3]. De nouvelles straté-
Tirés à part : D. Hober
abc
Ann Biol Clin 2005 ; 63 (6) : 581-8
Ann Biol Clin, vol. 63, n° 6, novembre-décembre 2005 581
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

gies thérapeutiques sont donc nécessaires. Parmi celles-ci,
l’immunothérapie spécifique ou vaccination thérapeutique
constitue une approche innovante et additionnelle pour le
traitement de l’infection par le VIH.
Structure et physiopathologie du VIH
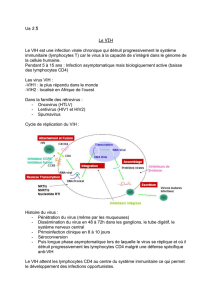
Structure du VIH (figure 1)
L’enveloppe virale formée d’une double couche lipidique
d’origine cellulaire, contient deux glycoprotéines asso-
ciées : la gp120 extra-membranaire et la gp41 transmem-
branaire. La face interne de l’enveloppe est tapissée d’une
couche protéique virale appelée matrice ou p17. La cap-
side contient deux copies identiques d’ARN monocaté-
naire, ainsi que les enzymes virales : la transcriptase
inverse, l’intégrase et la protéase. Elle contient également
deux protéines : la p24 protéine majeure de capside et la
p7 protéine de nucléocapside [6].
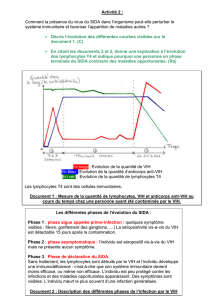
Organisation génomique du VIH
Le génome du VIH (figure 2) comporte trois gènes de
structure gag, pol et env — codant respectivement les
protéines internes, les trois enzymes virales et les glyco-
protéines d’enveloppe — et 6 gènes régulateurs de la
réplication virale : tat, rev, vif, nef, vpr et vpu [6].
Variabilité du VIH
La variabilité du virus est très grande. Elle résulte des
erreurs de copies effectuées par la transcriptase inverse
lors de la réplication. Le virus existe chez les patients sous
forme d’une multitude de variants (quasi-espèce). La
variabilité n’est pas la même tout le long du génome. La
région env qui code la gp120 et qui contient un épitope
majeur de neutralisation ainsi que des déterminants de
réponses immunes cytotoxiques est la plus variable. De
plus, il existe plusieurs sous-types du VIH-1 ainsi que des
formes recombinantes [7, 8].
Pathogenèse de l’infection
L’infection par le VIH est une infection virale chronique
avec une réplication continue au niveau du tissu lym-
phoïde. Le principal récepteur du VIH est la molécule
CD4 présente sur les lymphocytes T CD4+ mais égale-
ment à la surface d’autres cellules : monocytes sanguins,
macrophages tissulaires et cellules dendritiques. La
primo-infection s’accompagne d’une virémie massive et
d’une diminution du nombre de lymphocytes T CD4+. La
dissémination du virus dans les compartiments cellulaires
cibles, lymphatiques et neurologiques en particulier, est
très rapide. Dans plus de la moitié des cas, des manifesta-
tions cliniques accompagnent la primo-infection. Il peut
s’agir de manifestations graves, notamment neurologi-
ques : méningites et surtout encéphalites [9]. L’apparition
d’une forte réponse des lymphocytes cytotoxiques T
CD8+ permet de contrôler progressivement cette virémie
[10]. La primo-infection est suivie d’une phase dite de
latence clinique. Le virus persiste néanmoins à l’état
infectieux dans les organes lymphoïdes (réservoirs) où il
continue de se répliquer de manière chronique. Une dété-
rioration qualitative du système immunitaire s’installe et
évolue vers un déficit quantitatif sévère responsable des
manifestations cliniques du sida [10].
Mécanismes immunologiques
impliqués dans l’infection à VIH
Déficit immunitaire induit par le VIH
L’infection par le VIH est associée à des anomalies immu-
nologiques qui apparaissent précocement dans l’histoire
naturelle de l’infection [11]. Ainsi le déficit quantitatif des
lymphocytes T CD4+, lié à l’effet direct du virus et à la
destruction des cellules infectées par le système immuni-
taire, est précédé par l’existence d’anomalies fonctionnel-
les des lymphocytes T CD4+, T CD8+, des monocytes et
des macrophages. L’infection par le VIH s’accompagne
d’une dérégulation du réseau des cytokines et d’une modi-
fication de leur production. Cette dérégulation joue un rôle
dans l’activation chronique du système immunitaire obser-
vée pendant toute l’évolution de l’infection et favorise
l’apoptose (mort cellulaire programmée) et l’anergie des
lymphocytes T [4, 11].
Restauration partielle du système immunitaire
sous traitement antirétroviral
L’administration d’une trithérapie antirétrovirale efficace
est associée à une augmentation significative du taux des
P17 matrice
P24 capside
P7 nucléocapside
Intégrase
RT / TI
Transcriptase
inverse
Protéase
Gp120
Gp41
90 à 120 nm
ARN
Capside
Enveloppe
Figure 1. Structure du VIH.
revue générale
Ann Biol Clin, vol. 63, n° 6, novembre-décembre 2005582
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

lymphocytes T CD4+ mémoires et CD4+ naïfs et à une
restauration progressive et lente des fonctions lymphocy-
taires. La reconstitution immunitaire est marquée par une
diminution des signes d’hyperactivation des lymphocytes
T CD8+ et des lymphocytes T CD4+ ; la restauration de la
réponse proliférative des lymphocytes aux antigènes de
rappel et par une réduction de l’apoptose des lymphocytes
T [4, 11]. Toutefois, sous traitement antirétroviral, la
reconstitution immunitaire n’est jamais totale [11]. Le
traitement permet une restauration des réponses immunes
vis-à-vis des agents infectieux opportunistes mais les
réponses immunes spécifiques anti-VIH sont rarement res-
taurées lorsque l’infection est traitée au stade chronique.
Quand le traitement de l’infection par le VIH est instauré
durant la phase aiguë, les lymphocytes T CD4+ auxiliaires
spécifiques du VIH sont préservés de la destruction mais
le développement des réponses CD8+ est limité [3].
Vaccin thérapeutique
contre l’infection par le VIH
Définition
Le vaccin thérapeutique consiste à utiliser des prépara-
tions vaccinales pour induire ou amplifier les réponses
immunitaires anti-VIH chez des patients infectés par le
VIH. Il s’agit d’une immunothérapie spécifique. Il est
conceptuellement différent du vaccin préventif qui a
comme objectif d’empêcher l’infection [12].
Rationnel
Le concept d’immunisation thérapeutique comme traite-
ment du VIH a été proposé pour la première fois par Jonas
Salk [13]. Ce dernier a suggéré que la longue période
séparant la primo-infection VIH-1 et le développement du
sida était due à une réponse immunitaire spécifique qui, si
elle était renforcée réduirait la charge virale et ralentirait
la progression de la maladie.
En effet, un certain nombre d’observations cliniques et
expérimentales suggèrent que le système immunitaire est
capable de contrôler efficacement la réplication virale.
Durant la primo-infection, la baisse de la charge virale est
contemporaine de l’apparition d’une réponse immunitaire
spécifique liée aux lymphocytes T CD4+ et CD8+ anti-
VIH. Ultérieurement, en l’absence de traitement antirétro-
viral, la réactivité anti-VIH des lymphocytes T CD4+
n’est plus détectée chez la majorité des patients. Elle per-
siste cependant chez les patients asymptomatiques à long
terme où son rôle est évoqué dans le contrôle de la réplica-
tion virale. La persistance des lymphocytes T CD8+ anti-
VIH à réactivité polyépitopique est également associée à
l’absence d’évolution du déficit immunitaire chez les
patients non traités [3, 8]. L’intensité de la réponse T
cytotoxique est plus forte chez les sujets asymptomatiques
à long terme que chez les sujets qui progressent vers le
déficit immunitaire. Des facteurs génétiques semblent
également jouer un rôle dans ce phénomène de « non pro-
gression » [3, 14].
Chez les sujets non infectés malgré une exposition répétée
sur plusieurs années au VIH-1, des lymphocytes T cyto-
toxiques anti-VIH circulants ou muqueux ont été mis en
évidence [8, 11, 15]. Une délétion homozygote dans le
gène codant un corécepteur du VIH-1 (CCR5) a été
retrouvée chez des sujets exposés et non infectés d’origine
caucasienne [16]. Les défenses antivirales innées pour-
raient également être impliquées dans la résistance à
l’infection [17].
Au cours des essais d’interruptions thérapeutiques, l’appa-
rition de réponses cellulaires spécifiques du VIH est obser-
vée lors de la remontée de la réplication virale [18]. Enfin,
la déplétion des lymphocytes T CD8+ chez les macaques
infectés par le virus de l’immunodéficience simienne
aboutit à une réplication virale non contrôlée et une évolu-
tion rapide vers le déficit immunitaire [4].
Objectifs immunologiques
La vaccination thérapeutique a pour but d’amplifier ou de
renforcer les réponses immunitaires spécifiques anti-VIH
chez des personnes déjà infectées afin de contrôler la
réplication virale [18, 19]. Elle permettrait également
d’apporter des capacités immunitaires additionnelles pour
couvrir certains épitopes qui ont été négligés par l’hôte
infecté, ou qui sont apparus après mutation du virus [20].
LTR
Gag
Pol Vif Tat
Vpr Rev
Vpu Tat
Rev
Nef
LTR
Env
Figure 2. Organisation génomique du VIH. L’organisation du génome viral comporte des gènes de structure ainsi que différents gènes
de régulations. Le gène gag code les protéines internes : protéine de matrice, protéine de la capside et protéine associée à l’ARN. Le
gène pol code différentes enzymes virales : protéase, transcriptase inverse et intégrase. Le gène env code les glycoprotéines externes.
Aux extrémités du génome viral, se trouvent des séquences répétitives inversées (LTR ou long terminal repeats) contenant l’ensemble
des signaux nécessaires à la transcription et l’intégration du virus.
Vaccination thérapeutique et VIH-1
Ann Biol Clin, vol. 63, n° 6, novembre-décembre 2005 583
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

Objectifs cliniques
Les objectifs cliniques de la vaccination thérapeutique
sont multiples. Ils visent à retarder la progression de
l’infection VIH ou l’initiation du traitement antirétroviral
(notamment dans les pays à faibles ressources économi-
ques) ; améliorer l’efficacité du traitement antirétroviral et
permettre une interruption thérapeutique. Le but ultime est
l’arrêt définitif du traitement antirétroviral. Ceci permet-
trait une réduction des coûts, de la toxicité et le repos des
patients [18-20].
Obstacles au développement d’un vaccin anti-VIH
De nombreux obstacles techniques et biologiques compli-
quent la mise au point du vaccin thérapeutique (tableau 1)
[7, 8]. Les épitopes cibles des anticorps neutralisants des
souches de virus sauvages sont des épitopes conforma-
tionnels qui sont très peu immunogènes et qui sont mas-
qués [21]. De plus, la boucle V3 de la glycoprotéine de
surface gp120, considérée comme contenant l’épitope
majeure de neutralisation, est d’une variabilité extrême.
Par ailleurs, il n’est pas exclu que la vaccination puisse
induire des anticorps pouvant exacerber l’infectiosité. Il a
été suggéré que des anticorps facilitants pouvaient favori-
ser la pénétration du virus dans une cellule cible ou même
permettre son accès à des cellules dépourvues de récep-
teurs spécifiques du virus [8, 22].
En ce qui concerne les réponses cellulaires, les lymphocy-
tes T ne reconnaissent l’antigène que sous la forme de
peptides apprêtés, présentés en association avec la molé-
cule du complexe d’histocompatibilité (CMH). Vu la
grande diversité des haplotypes des antigènes du CMH au
sein de la population humaine, le vaccin doit contenir un
large répertoire d’épitopes T pour induire une réponse
efficace chez tous les vaccinés [21].
Enfin, les quelques modèles animaux disponibles, très
onéreux et éloignés du modèle humain ne permettent de
réaliser des essais que sur un petit nombre d’animaux
avant de passer à des essais humains. Le meilleur modèle
est celui du macaque infecté par un virus hybride SHIV
(simian-human imunodeficiency virus) qui présente une
maladie proche du sida [7, 8].
Les antigènes vaccinaux
Les protéines d’enveloppe synthétisées par la région env
sont des inducteurs importants d’anticorps neutralisants
anti-VIH. L’hypervariabilité de la boucle V3 de la gp120
explique le faible pouvoir neutralisant des anticorps. Les
protéines Gag (p17, p24), relativement conservées, sont
une cible préférentielle de la réponse CTL
(cytotoxic T lymphocytes) mais l’échappement immuni-
taire est possible. Les protéines des gènes régulateurs (nef,
tat,rev essentiellement) semblent être d’excellents immu-
nogènes. Elles sont de plus en plus utilisées [5, 8, 20]. La
protéine Tat est essentielle pour le cycle viral du VIH-1.
Elle est exprimée très tôt après l’entrée du virus dans la
cellule. Une réponse humorale et cellulaire vis-à-vis de
Tat serait liée à une lente progression de la maladie chez
l’homme et le singe [7].
Le vaccin idéal
Le vaccin idéal doit permettre de déclencher une réponse à
la fois humorale et cellulaire (tableau 2) [8]. Les anticorps
sont actifs contre les virus libres et pourraient inhiber les
protéines toxiques secrétées durant la maladie (Nef, Tat)
[20, 23]. Les réponses des lymphocytes T CD8+ cytotoxi-
ques (CTL) sont très souhaitées car elles peuvent réduire
la production virale en éliminant les cellules infectées. Les
lymphocytes T CD8+ exercent ce contrôle de la réplica-
tion du VIH via une activité cytotoxique et/ou la produc-
tion de cytokines et chimiokines ayant un effet suppres-
seur de la réplication virale [11]. Les lymphocytes CD4+
T auxiliaires type 1 (Th1) sont essentiels pour une activité
CTL (voir illustration dans [24]). Actuellement, en raison
des difficultés à générer des anticorps capables de neutra-
liser les isolats primaires, l’accent a été mis sur les répon-
ses cellulaires [8, 19, 20]. Il n’est pas exclu que des anti-
corps, pouvant exacerber l’infectiosité, puissent
apparaître.
Méthodes d’évaluation
de l’immunogénicité des vaccins
L’immunogénicité vaccinale est mesurée par une variété
de paramètres immunologiques qui tentent d’évaluer les
réponses anticorps neutralisantes et cytotoxiques. On peut
citer le test de lyse des cellules infectées, la technique des
tétramères HLA, le test ELISpot interféron gamma (quan-
tification des lymphocytes T spécifiques d’un antigène par
la mesure de la production de l’interféron gamma), l’étude
de la prolifération des lymphocytes T CD4+ en présence
d’un antigène, la détection des anticorps neutralisants...
[3, 7, 18].
Ces différents tests ne donnent qu’une image partielle de
la réponse immune. Des tests complémentaires devront
être utilisés après validation et standardisation, notamment
Tableau 1. Obstacles au développement d’un vaccin anti-VIH.
- Atteinte précoce du système immunitaire
- Infection chronique latente échappant à la réponse immune
- Variabilité extrême du virus
- Médiocre immunogénicité des antigènes conservés
- Résistance à la séroneutralisation des isolats primaires
- Méconnaissance des marqueurs de protection
- Insuffisance des modèles animaux
revue générale
Ann Biol Clin, vol. 63, n° 6, novembre-décembre 2005584
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.

la mesure de l’IL-2, la mesure de la sécrétion de la perfo-
rine et de la Granzyme B [7].
Les marqueurs d’efficacité clinique des vaccins
De nombreux vaccins sont actuellement testés. Leur effi-
cacité clinique est évaluée sur la durée de l’interruption
thérapeutique qu’ils permettent [2]. L’efficacité immuno-
virologique est évaluée par la charge virale et le dosage
des lymphocytes T CD4+ [3].
Les candidats vaccins
Virus entier inactivé (Remune
®
)
Il s’agit d’un vaccin constitué de particules entières du
VIH-1 délétées de leur gp120 de surface. Les essais de
phase I ont montré que Remune
®
induisait une réaction
d’hypersensibilité retardée anti-VIH et augmentait le titre
des anticorps anti-p24 mais un essai randomisé de phase
III fût interrompu car on n’a pas pu montrer de différences
entre les groupes traités et non traités en terme de progres-
sion clinique (maladies opportunistes) et de décès. L’ana-
lyse des objectifs secondaires montre néanmoins un taux
de lymphocytes T CD4+ et une réponse lympho-
proliférative plus élevés dans le groupe des sujets vaccinés
[5].
Protéines recombinantes
Il s’agit principalement des glycoprotéines d’enveloppe
recombinantes gp120, gp41 ou de leur précurseur gp160,
cibles de la plupart des anticorps neutralisants. Les résul-
tats obtenus dans un essai de phase I montrent que l’utili-
sation de ces protéines est bien tolérée et permet d’obtenir
des réponses immunitaires humorales ou lymphocytaires.
Néanmoins, l’évaluation de ces protéines dans des essais
randomisés et contrôlés de phase II n’a pas montré d’effi-
cacité de la vaccination en termes de bénéfices cliniques
[5, 7].
Les pseudo-virions
Les pseudo-virions ou pseudo-particules virales (virale
like particules ou VLP) contiennent tous les composants
protéiques du virus sauf le génome. Des particules recom-
binantes Ty-VLP fabriquées à partir d’un transposon de
levures et contenant une protéine de fusion gag p17/p24 a
permis d’obtenir une réponse immune humorale et cellu-
laire anti-p24 spécifiques chez les primates non humains.
Cependant, durant de multiples essais de phase II, ces
vaccins n’ont pas montré de bénéfice en termes clinique
ou de marqueurs biologiques [5].
Peptides ou lipopeptides
Ils reposent sur la synthèse chimique de peptides. Le but
est de susciter une réponse immune au niveau d’une
séquence virale polypeptidique d’intérêt immunologique
majeur comme par exemple un épitope CTL dominant. On
tend à utiliser un mélange de peptides contenant chacun
de multiples épitopes (gag, boucle V3...). La liaison cova-
lente de ces peptides à des lipides a permis d’augmenter
leur immunogénicité et en particulier d’améliorer considé-
rablement la réponse CTL [7, 20].
Vecteurs recombinants
Ils sont construits à partir de bactéries ou de virus inoffen-
sifs pour l’homme dans lesquels sont insérés des gènes du
VIH (généralement pol,gag et env). Ces vecteurs entrent
dans les cellules et expriment lors de leur réplication une
ou plusieurs protéines virales. Ces protéines virales sont
présentées sous leur forme native au système immunitaire.
Un certain nombre de vecteurs viraux sont actuellement
testés : Canarypox (Alvac), adénovirus type 5 (Ad5),
adeno-associated virus (AAV), modified vaccinia ankara
Tableau 2. Différentes approches pour le développement d’un vaccin anti-VIH. (D’après [8]).
Avantages Inconvénients et obstacles
Vaccin à
réponse CTL
Reconnaissance des cellules infectées
Épitopes linéaires multiples
Élimination de la production virale
Effet possible sur le réservoir latent
Nécessité de lymphocytes mémoires de
longue vie
Incapacité de reconnaissance du virus en
l’absence de la molécule CMH capable de
se lier au peptide viral
Régulation négative des molécules CMH par
le virus
Échappement immunitaire possible
Vaccin à
réponse
anticorps
Neutralisation du virus
Capacité d’empêcher une nouvelle infection
Activation de la réponse inflammatoire
(système du complèment, neutrophiles,
monocytes)
Incapacité à générer une réponse
neutralisante large
Hypervariabilité des épitopes de
neutralisation
Anticorps pouvant amplifier l’infectiosité
Épitopes conformationnels
Vaccination thérapeutique et VIH-1
Ann Biol Clin, vol. 63, n° 6, novembre-décembre 2005 585
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 24/05/2017.
 6
7
8
6
7
8
1
/
8
100%