Les crises d`absence - John Libbey Eurotext

doi: 10.1684/epi.2010.0287
Les crises d’absence :
des pointe-ondes généralisées
initiées par un foyer cortical
Pierre-Olivier Polack
1,2
, Stéphane Charpier
1
1
UPMC/Inserm UMR-S 975, CNRS UMR 7225, Centre de recherche de l’institut du cerveau
et de la moelle épinière, hôpital Pitié-Salpêtrière, 75013 Paris, France
2
Department of Neuroscience–School of Medicine, University of Pennsylvania, 121, Anatomy-
Chemistry Building, Philadelphia, PA 19104, États-Unis
Résumé. Les crises d’épilepsie sont classées par l’International League Against Epilepsy en deux grands grou-
pes : les crises partielles, initiées par une zone épileptogène, et les crises généralisées. Les crises d’absence sont des
crises généralisées dont les décharges de pointe-ondes (DPO) sont exprimées au sein de la boucle thalamocorticale.
L’origine, corticale ou thalamique, des paroxysmes électriques associés aux crises d’absence a été l’objet d’intenses
controverses. Confirmant de récentes données électrocliniques obtenues chez l’homme, indiquant que les absences
sont initiées à partir de zones corticales discrètes, des études réalisées sur le rat WAG/Rij et le GAERS (Genetic
Absence Epilepsy Rat from Strasbourg), deux modèles génétiques présentant une forte homologie avec la pathologie
humaine, ont révélé l’existence d’un foyer cortical à l’origine des DPO. Nous avons précisé les mécanismes de
déclenchement des crises en identifiant chez le GAERS une sous-population de neurones corticaux, hyperactifs et à
décharges ictogènes précoces, initiant les DPO dans la boucle corticothalamique. L’ensemble de ces données expé-
rimentales et cliniques supporte fortement l’existence d’un foyer cortical à l’origine des crises d’absence, remettant
ainsi en cause les classifications électrocliniques et étiologiques « classiques » de l’épilepsie-absences.
Mots clés :épilepsie-absences,cortex,thalamus,modèle animal,GAERS,foyer cortical
Abstract. Absence seizures: generalized spike-and-wave discharges initiated from a cortical focus
Epileptic seizures are classified by the International League Against Epilepsy into two main groups: partial seizures,
initiated from an epileptogenic zone, and generalized seizures. Absence seizures are part of generalized seizures, with
spike-and-wave discharges (SWDs) occurring within the thalamo-cortical loop. The origin, cortical or thalamic, of
electrical paroxysms associated with absence seizures has been subject of fervent controversies. Consistent with recent
electro-clinic findings obtained in human patients, indicating that absences are initiated from discrete cortical zones,
studies carried out on WAG/Rij rats and GAERS (Genetic Absence Epilepsy Rat from Strasbourg), two genetic models
exhibiting a strong homology with the human pathology, revealed the existence of a cortical focus triggering the
SWDs. We specified the mechanisms of seizure initiation by identifying in the GAERS a subpopulation of cortical
neurons, hyperactive and showing early ictogenic discharges, from which the SWDs emerge and are secondarily
propagated within the cortico-thalamic loop. Altogether, these experimental and clinical findings strongly support the
existence of a cortical focus initiating absence seizures, leading to a re-evaluation of the “classical”electro-clinic and
etiologic classification of absence epilepsy.
Key words:absence epilepsy,cerebral cortex,thalamus,animal model,GAERS,cortical focus
Épilepsie et recherche
Épilepsie et recherche
Épilepsies 2010 ; 22 (1) : 33-41
Tirés à part :
P.-O. Polack
Épilepsies, vol. 22, n° 1, janvier-février-mars 2010
33
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

Introduction
La boucle thalamocorticale est le siège des décharges
de pointe-ondes
L’International League Against Epilepsy, dans sa classification
des crises d’épilepsie (ILAE, 1981) distingue deux grands
groupes, les crises généralisées et les crises partielles. Les crises
généralisées sont définies par la survenue de décharges
paroxystiques apparemment simultanément dans les deux
hémisphères. Les crises partielles ou focales, sont caractérisées
par des décharges épileptiques dans une région délimitée du
cerveau, le plus souvent dans le cortex cérébral, appelée
« zone épileptogène » ou « foyer épileptique » (ILAE, 2003).
La distinction entre crises partielles et crises généralisées se
base essentiellement sur des critères symptomatiques et élec-
troencéphalographiques (EEG), mais ne permet en rien de
préciser les processus ictogènes sous-jacents. En effet, il est
difficilement concevable que l’initiation d’une crise d’épilepsie
généralisée résulte d’une synchronisation paroxystique, spon-
tanée et immédiate, de la quasi-totalité des neurones de
l’ensemble du cortex. Il a donc été supposé l’existence de terri-
toires épileptogènes restreints à l’origine des crises généralisées,
lesquels demeurent inaccessibles aux moyens d’investigation
clinique habituels.
Les crises d’absence typiques font partie de ces crises épi-
leptiques généralisées dont les mécanismes physiopathologi-
ques restent mal connus. Elles sont définies cliniquement
comme des altérations transitoires de la conscience associées à
la présence de décharges de pointe-ondes (DPO) bilatérales,
synchrones et symétriques dans l’électroencéphalogramme
(EEG). Les DPO surviennent et disparaissent brutalement sur
une activité EEG de fond normale et sont apparemment
d’emblée généralisées à l’ensemble de la surface corticale.
Le substrat neuroanatomique des crises d’absence est la
boucle thalamocorticale. La première démonstration directe
de la présence d’une activité épileptique dans le thalamus
lors des crises d’absence a été obtenue en 1953, chez une
patiente de cinq ans, où des DPO ont pu être enregistrées
simultanément dans les EEG corticaux et thalamiques lors de
la crise (Williams, 1953). L’existence d’un désordre fonction-
nel dans le cortex cérébral et le thalamus lors des DPO a été
confirmée par des études combinant imagerie tomographique
par émission de positons (SPECT) ou imagerie par résonance
magnétique fonctionnelle (IRMf) et par enregistrements
d’EEG, révélant une hyperactivité métabolique dans ces deux
structures lors de crises d’absence (Iannetti et al., 2001 ;
Aghakhani et al., 2004 ; Labate et al., 2005).
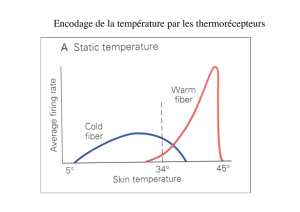
La boucle thalamocorticale (figure 1) est composée de neu-
rones corticothalamiques excitateurs (glutamatergiques), locali-
sés dans les couches profondes du cortex (couches V et VI),
connectant les neurones thalamocorticaux qui, en retour, exci-
tent par des synapses glutamatergiques les neurones corticaux.
Les terminaisons axonales des neurones corticothalamiques de
la couche VI, ainsi que celles des neurones thalamocorticaux
contactent également les neurones gabaergiques du noyau
réticulaire du thalamus (nRT), lesquels inhibent en retour les
neurones thalamocorticaux (Jones, 2002). Les circuits thalamo-
corticaux sont donc composés d’une large boucle thalamocor-
ticale excitatrice et d’une boucle locale thalamothalamique
constituée de connexions excitatrices et inhibitrices récipro-
ques. Cette configuration anatomique, de type « cyclique », a
pour conséquence une interdépendance des activités thalami-
ques et corticales et rend délicate la mise en évidence d’un site
d’initiation des crises d’absence.
Synopsis de la revue
Dans cette revue, nous présenterons les différentes hypo-
thèses d’initiation des crises d’absence, au niveau thalamique
et cortical, proposées sur la base de recherches cliniques et
fondamentales. Nous détaillerons les travaux que nous
avons réalisés et qui ont permis de mettre en évidence le
site cortical et les neurones ictogènes spécifiques, à partir des-
quels les crises sont initiées chez les GAERS (Genetic Absence
Epilepsy Rats from Strasbourg), le modèle génétique le mieux
validédel’épilepsie-absences. Nous discuterons du rôle des
différents compartiments de la boucle thalamocorticale dans
l’initiation, la propagation et la terminaison des crises
d’absence, ainsi que de l’action antiépileptique de l’éthosuxi-
mide (ETX), traitement de première intention chez l’homme,
sur les neurones ictogènes du foyer cortical. Nous conclurons
en proposant une extrapolation de nos résultats et hypothè-
ses aux crises d’absence humaines, conduisant à reconsidérer
la classification « classique » des épilepsies-absences.
Origines des crises d’absence :
du thalamus au cortex
Les DPO apparaissant de manière quasi simultanée sur de
vastes territoires corticaux, il a initialement été proposé que
lescrisesd’absence ont une origine extrinsèque au cortex, et
en particulier sous-corticale (Jasper et Kershman, 1941). À la
même époque, Morison et Dempsey décrivent les noyaux
thalamiques intralaminaires qui présentent des projections
diffuses vers le cortex, contrairement aux noyaux thalami-
ques de relais qui focalisent leurs terminaisons axonales vers
un territoire cortical spécifique (Morison et Dempsey, 1942).
Étant donné leurs propriétés anatomofonctionnelles, les
noyaux intralaminaires du thalamus sont devenus un
« bon » candidat pour un « pacemaker extracortical » capable
d’activer simultanément de vastes territoires corticaux. Cette
hypothèse était également supportée par les stimulations
électriques des noyaux intralaminaires, chez le chat et chez
le singe, induisant un arrêt comportemental associé à une
survenue de DPO bilatérales et synchrones à trois cycles par
seconde (Jasper et Droogleever Fortuyn, 1947). L’hypothèse
thalamique fut reprise et développée dans les années 1990
par György Buzsáki et David McCormick, (McCormick et
Contreras, 2001) montrant in vivo (Buzsaki, 1991) et in vitro
P.-O. Polack, S. Charpier
Épilepsies, vol. 22, n° 1, janvier-février-mars 2010 34
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

(Bal et al., 1995a ; Bal et al., 1995b) que les neurones du nRT
et les neurones thalamiques de relais peuvent former un
« système oscillateur » capable de maintenir une activité ryth-
mique et synchrone à 3 Hz dans la boucle thalamocorticale
(figure 1) (McCormick et Contreras, 2001).
Bien que ces données puissent théoriquement expliquer la
soudaine généralisation des crises d’absence à partir d’un dys-
fonctionnement thalamique, elles ont été remises en cause par
de nombreuses expériences et observations cliniques démon-
trant le rôle crucial du néocortex dans l’initiation des absences.
L’analyse rigoureuse de tracés d’EEG de malades atteints d’épi-
lepsies généralisées (Gibbs et Gibbs, 1952), ainsi que des expé-
riences menées sur des patients souffrant d’épilepsie-absences
(Bennett, 1953), ont conduit Pierre Gloor à proposer une ori-
gine « corticoréticulaire » aux épilepsies-absences, c’est-à-dire
procédant d’interactions complexes entre le cortex et le nRT
(figure 1). Il reproduisit chez des patients et chez des chats les
expériences de Bennett montrant que l’injection de produits
proconvulsivants, tel que le pentylenetetrazol, dans l’artère
carotide (qui perfuse le cortex), générait des DPO généralisées,
alors que la même injection réalisée dans l’artère vertébrale
(qui perfuse le diencéphale et le tronc encéphalique) ne pro-
duisait aucun effet (Bennett, 1953 ; Gloor, 1968 ; Gloor,
1969 ; Meeren et al., 2005). D’autres cliniciens et neurophysio-
logistes proposèrent sur la base d’observations électrocliniques,
une initiation corticale des crises au niveau de foyers diffus
localisés dans les cortex frontaux ou mésofrontaux, le thala-
mus participant à la généralisation des paroxysmes en relayant
l’activité rythmique d’origine corticale (figure 1) (Bancaud,
1969 ; Niedermeyer, 1972 ; Luders et al., 1984 ; Niedermeyer,
1996). Ces propositions furent supportées par les travaux expé-
rimentaux de Pierre Gloor sur le modèle « chat pénicilline »
montrant que l’application diffuse sur la surface corticale de
pénicilline, un antagoniste faible des récepteurs GABA(A),
générait des DPO, alors qu’une injection de cette substance
proépileptique au niveau thalamique restait sans effet (Gloor
et al., 1990). Sur la base de ces dernières observations, Gloor
et al. conclurent que les DPO dans la boucle thalamocorticale
sont générées à partir du cortex quand celui-ci se trouve dans
un état d’hyperexcitabilité, le thalamus jouant un rôle mal
défini mais néanmoins indispensable dans le maintien des
activités paroxystiques synchrones.
Théorie du foyer cortical
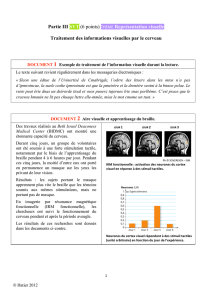
En 2002, les équipes de Fernando Lopes da Silva et Gilles
van Luijtelaar analysèrent l’association non linéaire entre des
signaux EEG enregistrés simultanément dans le cortex et le
thalamus au cours de crises d’absence spontanées chez des
rats WAG/Rij, un modèle génétique d’épilepsie-absences
(figure 2A). Ils montrèrent par cette approche mathématique
sophistiquée l’existence d’un décalage de quelques millisecon-
des entre les DPO enregistrées dans une région particulière du
cortex somatosensoriel (l’aire somatosensorielle périorale) et
celles d’autres territoires thalamiques et corticaux (figure 2B)
Meeren et al., 2002). Ce délai entre les activités épileptiques
spatialement distribuées suggérait que chaque pointe-onde est
initiée par le cortex somatosensoriel périoral puis se propage à
la vitesse de 2 m/s aux autres cortex et au thalamus (Meeren
et al., 2002).
Pour déterminer si l’existence d’un foyer cortical est une
spécificité des rats WAG/Rij ou bien une caractéristique
commune à l’ensemble des modèles génétiques d’épilepsie-
absences, nous avons réalisé in vivo des enregistrements
simultanés des potentiels de champs locaux de différents ter-
ritoires cérébraux, incluant les cortex moteurs et somatosen-
soriels et diverses régions thalamiques chez un autre modèle
d’absence, les GAERS (figure 2C). Ces enregistrements ont
révélé que les DPO enregistrées au cours de la crise dans le
cortex somatosensoriel périoral précèdent de quelques milli-
secondes celles enregistrées dans les autres régions corticales
et thalamiques, suggérant que les activités paroxystiques se
propagent à partir de ce territoire vers les autres régions corti-
cales et thalamiques (figure 2C2) (Polack et al., 2007 ; Polack
et al., 2009a). Ces enregistrements multi-sites ont également
Cortex
Hypothèse corticale
Hypothèse
cortico-réticulaire
hypothèse thalamique
Noyau de relais nRT
Neurone Synapse
excitateur
(Glutamate)
inhibiteur
(GABA)
II/III
IV
V
VI
Figure 1. Représentation schématique de la boucle thalamocorti-
cale et des différentes hypothèses sur le déclenchement des crises
d’épilepsie-absences. Les neurones excitateurs (en noirs) utilisent le
glutamate comme neurotransmetteur. Les neurones du noyau réti-
culaire du thalamus (nRT) sont gabaergiques (cercles). Les interneu-
rones inhibiteurs localisés dans les différentes couches corticales
ont été omis. Les régions cérébrales concernant les trois théories
(« corticale », « thalamique » et « corticoréticulaire ») sur le site de
déclenchement des crises sont indiquées par les crochets.
Les crises d’absence sont-elles des crises focales ?
Épilepsies, vol. 22, n° 1, janvier-février-mars 2010
35
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

montré que les activités paroxystiques pouvaient survenir
dans le cortex somatosensoriel périoral du GAERS quelques
secondes avant qu’une activité épileptique ne soit détectée
dans les autres régions corticales ou thalamiques (figure 2C1)
(Polack et al., 2007 ; Polack et al., 2009a). Cette activité ictale
locale pouvait également rester localisée au seul cortex soma-
tosensoriel (figure 2D) (Polack et al., 2007). Les GAERS,
comme les rats WAG/Rij, présentent donc une zone, ou
foyer, épileptogène localisé dans le cortex somatosensoriel
périoral, initiant les activités paroxystiques et les propageant
ensuite à l’ensemble du cortex et du thalamus.
Les neurones des couches profondes
du cortex somatosensoriel périoral
des GAERS présentent une hyperactivité
spécifique
Une fois localisée la région cérébrale initiant les activités
paroxystiques chez les GAERS, nous avons cherché à détermi-
ner les propriétés électriques et l’activité des neurones de ce ter-
ritoire au cours des crises d’absence. Nous avons enregistré
in vivo chez les GAERS l’activité intracellulaire des neurones
localisés dans les différentes couches du cortex somatosensoriel
périoral lors de la survenue spontanée des DPO (figure 3B). Nous
avons ensuite comparé cette activité à celle de neurones enre-
gistrés dans le cortex moteur des mêmes animaux et de neuro-
nes de la même région anatomique chez des animaux non
épileptiques (figure 3) (Polack et al., 2007). Au cours des crises,
les neurones des couches profondes (couches V et VI) se distin-
guent des autres neurones enregistrés par une forte décharge de
potentiels d’action qui précède dans le temps celles des neuro-
nes des couches plus superficielles (couche IV et couches II/III)
(figure 3C) et des neurones des couches profondes du cortex
moteur. Au cours des périodes interictales, les neurones des
couches profondes se distinguent également des autres popula-
tions neuronales par un potentiel de membrane significative-
ment plus dépolarisé et une décharge de potentiels d’action
plus élevée et plus régulière (figure 3A). Des enregistrements
des neurones des couches profondes du cortex somatosensoriel
périoral (région homotypique du foyer cortical du GAERS) de
rats non épileptiques ont démontré que cette hyperactivité est
spécifique des neurones des couches profondes de la région
initiant et propageant les pointe-ondes (figure 3A).
AB1
C1 C2 D
B2
7
2
3
4
5
6
7
latéral (mm)
654321
Rat WAG/Rij
5
mV
500 ms
H11
H12
H15
H16
H18
H19
H20
H21
012345
posterieur (mm)
bregma
antérieur (mm)
WAG/Rij (EEG)
GAERS (Potentiels de Champs Locaux)
cortex
somatosensoriel
1
2
3
4
5
6
7
cortex
moteur
thalamus
500 ms
C2 100 ms 1 sec
Figure 2. Mise en évidence d’un foyer cortical à l’origine des crises dans deux modèles animaux présentant des crises d’absence sponta-
nées. A) Position des électrodes d’EEG sur la surface corticale du rat à partir desquelles les résultats présentés en B ont été obtenus (les élec-
trodes intrathalamiques ont été omises) ; B) enregistrement EEG multi-unitaire d’une crise d’absence chez le rat WAG/Rij et localisation du
foyer cortical à partir de cet enregistrement. B1. Exemple d’enregistrements obtenus à partir de l’arrangement expérimental schématisé en
A. B2. Représentation topographique des foyers corticaux de huit rats représentés par des symboles différents (insert). Les symboles noirs
indiquent les sites corticaux dont l’activité ictale précède celle de tous les autres sites enregistrés. La zone délimitée par l’ovale correspond
au cortex somatosensoriel périoral ; C) enregistrement multi-unitaire des potentiels de champs locaux, corticaux et thalamiques, chez un
GAERS. C1. Les activités paroxystiques sont d’abord localisées au niveau du cortex somatosensoriel, puis se propagent au cortex moteur et
au thalamus. C2. Agrandissement de la succession de trois pointe-ondes indiquées par le cadre gris en C1. Le pic de négativité des pointes
enregistrées dans le cortex somatosensoriel précède celui des pointes enregistrées dans le cortex moteur et le thalamus ; D) enregistrement
chez le même animal d’une activité paroxystique de courte durée restant localisée au cortex somatosensoriel. B modifié d’après Meeren
et al. (2002).
P.-O. Polack, S. Charpier
Épilepsies, vol. 22, n° 1, janvier-février-mars 2010 36
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.

Lesneuronesdescouchesprofondesdufoyercortical
présentent également la propriété particulière de générer de
courtes activités ictales lorsqu’aucune activité épileptique
n’est visible dans l’EEG local.
Ces courtes activités oscillatoires paroxystiques peuvent
rester isolées ou donner naissance progressivement à la crise.
Les neurones hyperactifs des couches profondes du cortex
somatosensoriel périoral seraient donc capables d’initier des
activités paroxystiques et de soutenir la crise une fois celle-ci
généralisée (Polack et al., 2007).
Le cortex somatosensoriel périoral
des GAERS est nécessaire et suffisant
pour générer les DPO
Afin de préciser le caractère ictogène des neurones des cou-
ches profondes du cortex somatosensoriel périoral des GAERS,
il était nécessaire de tester la capacité de ces neurones à générer
de manière endogène des activités paroxystiques et à induire
les DPO à distance. Nous avons donc étudié les effets de leur
inactivation fonctionnelle sur la survenue des oscillations
paroxystiques locales et distantes. De plus, pour démontrer le
A
Intra
Intra
Intra
-58 mV
-64 mV
-63 mV
"Foyer" (couches V-VI) Foyer (couches V-VI)
GAERS (même rat)
Wistar
Contex moteur
EEG
EEG
EEG
B
C
Enregistrement intracellulaire
cortex moteur
Enregistrement intracellulaire
cortex somatosensoriel
EEG
cortex somatosensoriel
Foyer
couches V-VI
Foyer
couches II-III
Foyer
couches IV
Cortex moteur
couches V-VI
20 ms
20
mV
-64 mV
-79 mV
1 sec
Figure 3. Les neurones des couches profondes du cortex somatosensoriel périoral des GAERS sont hyperactifs et initient les décharges épi-
leptiques. A) Enregistrements de l’activité intracellulaire (traces du bas) d’un neurone des couches profondes (couches V-VI) du cortex soma-
tosensoriel périoral (foyer) et du cortex moteur chez le même GAERS, ainsi que d’un neurone du cortex somatosensoriel périoral d’un animal
non épileptique (wistar). L’activité intracellulaire est enregistrée simultanément avec l’activité d’EEG de surface du cortex somatosensoriel
(traces du haut). Les neurones du cortex somatosensoriel périoral des GAERS (panel du haut) présentent un potentiel membranaire plus
dépolarisé et une décharge de potentiels d’action plus élevée et plus régulière que les neurones du cortex moteur du même animal (panel
du milieu) et des neurones de la région homologue d’un animal non épileptique (panel du bas). La tête de flèche à gauche des enregistre-
ments intracellulaires indique le potentiel membranaire interictal moyen ; B) schéma du dispositif expérimental pour l’obtention des enregis-
trements présentés en A et C. Une électrode EEG est placée à la surface du cortex somatosensoriel périoral, les enregistrements
intracellulaires des neurones des colonnes corticales sous-jacentes sont réalisés chez des GAERS et des animaux non épileptiques (wistar).
Des enregistrements intracellulaires des neurones du cortex moteur sont ensuite réalisés chez le même animal ; C) superposition des enregis-
trements intracellulaires (traces du bas) d’un neurone des couches V-VI, des couches II-II et de la couche IV du cortex somatosensoriel pério-
ral ainsi que d’un neurone des couches V-VI du cortex moteur au cours d’une décharge de pointe-ondes enregistrée dans le cortex
somatosensoriel périoral (traces du haut). Les superpositions ont été réalisées en utilisant le pic de négativité de la pointe-onde comme réfé-
rence temporelle (0). Noter que la décharge de potentiels d’action des neurones des couches profondes du cortex somatosensoriel périoral
précède celle des neurones de la même région corticale plus superficiels ainsi que celle de neurones localisés dans les couches profondes
d’un cortex distant (moteur). C modifié d’après Polack et al. (2007).
Les crises d’absence sont-elles des crises focales ?
Épilepsies, vol. 22, n° 1, janvier-février-mars 2010
37
Copyright © 2017 John Libbey Eurotext. Downloaded by a robot coming from 88.99.165.207 on 24/05/2017.
 6
7
8
9
6
7
8
9
1
/
9
100%