Etude autoradiographique des premiers stades du

/. Embryol. exp. Morph.
Vol.
29, 3, pp. 585-600, 1973
585
Printed in
Great
Britain
Etude autoradiographique
des
premiers stades
du
developpement
de
Febauche
du membre anterieur chez deux especes
de
Cheloniens (Testudo graeca
L. et
Emys
orbicularis
L.)
Par
J.
VASSE1
Institut Pasteur, Sannois {Directeur: A. Raynaud)
SUMMARY
Autoradiographic study of
early
stages of the anterior limb-bud in
two species of
chelonian
embryos
Embryos
of
two
chelonian species
{Testudo graeca
L., Emys
orbicularis
L.) from 20-somites
stage
to
34-somites stage, fixed
2-4 h
after
an
injection
of
tritiated thymidine
or
uridine
or
leucine, then emulsion-exposed 12-14 days, showed the following results:
(1) When
the
labelling index
was
determined
in the
mesoderm
of the
somatopleure,
the
labelling index
in the
flank region (S15-S1G) remained
at
about 0-3
but the
labelling index
in
the prospective anterior limb region rose
to
0-4-0-5 when the processes
of
the somites S6-S13
reached
the
somatopleure. The somites seem
to
induce the proliferation
of
the somatopleure
and
the
transformation
of
the somatopleural cells
in
mesoblastic basophilic cells.
(2) The dilated extremities
of
the processes
of
somites S6-S12 proliferate cells which become
free from the somite and multiply. Thus, the somites S6-S12 participate materially
in
the con-
stitution
of the
mesoblastic blastema
of the
limb-bud.
(3)
In the
mesoblast, regionalizations appear
in
transverse histological sections
-
in
the
cranial-caudal direction.
(4)
An
apical epiblastic crest develops,
at
26-27-somites stage,
in the
area where
the
mesoblastic cells
are the
most abundant
and the
most active (between somites
S8 and Sn).
At 28-29-somites stage, this crest Anlage synthesizes very little
DNA; and at
34-somites
stage
the
synthesis
of
RNA
and
proteins also decreases.
These results
are
compared with results obtained
in
other reptiles
and
chick embryos,
in
the light
of
the theories
on
the role
of
the somites
and of
the apical crest
in
the development
of
the
anterior limb-bud.
INTRODUCTION
L'etude histologique des premiers stades du developpement du membre
anterieur chez l'embryon de Tortue mauresque (Testudo graeca L.) (Vasse &
Pieau, 1970) avait mis en evidence une heterogeneite de l'ebauche a ces stades
(structure differente du mesoblaste et de l'epiblaste suivant les niveaux somi-
1 Adresse de
Vauteur:
Service d'Embryologie experimentale, Institut Pasteur
20, rue des
Moulins, 95110 Sannois, France.
38-2

586 J. VASSE
tiques) et avait decrit les etapes de la formation de la crete apicale a partir d'un
epaississement localise de l'epiblaste. L'etude autoradiographique presente,
effectuee chez les embryons de Tortue mauresque et de Cistude d'Europe (Emys
orbicularis L.), a pour but de savoir comment se comportent les differentes
zones mises en evidence dans le mesoblaste et l'epiblaste (crete apicale en parti-
culier) en ce qui concerne les syntheses d'ADN, d'ARN et de proteines et de
suivre, si possible, revolution de ces syntheses.
La description se rapportera aux deux especes, les resultats etant tres com-
parables; dans le cas ou il existe des differences, l'espece sera mentionnee.
MATERIELS ET METHODES
Deux especes de Tortue ont ete utilisees: la Tortue mauresque (Testudo
graeca L.) originaire de Tunisie, elevee au laboratoire, et la Cistude d'Europe
(Emys orbicularis L.) provenant des etangs de Brenne (France). Les ceufs ont
ete pondus en juin-juillet dans les terrariums exterieurs avec des abris vitres
servant de refuge aux animaux la nuit ou par mauvais temps. Une ponte na-
turelle de Cistude trouvee dans le sol a ete egalement utilisee (aucune difference
avec les autres pontes n'a ete observee). Les oeufs ont ete mis en incubation sur
du coton remplissant un cristallisoir presque entierement recouvert d'une plaque
de verre de maniere a ne laisser qu'un petit espace pour les oeufs entre le coton
et la plaque de verre. Le coton est legerement humecte d'eau distillee dans la
partie inferieure du cristallisoir. Les temperatures d'incubation ont ete 25 et
30 °C. La vitesse du developpement varie avec la temperature d'incubation:
l'embryon de Cistude par exemple forme en 24 h 5-6 somites supplementaires
a 30 °C, 4 somites a 25 °C. Les stades seront exprimes en nombres de paires de
somites. Cinq stades seront distingues en ce qui concerne Pebauche du membre
anterieur. Le developpement
des
embryons des stades
1
a
3
est comparable a celui
du stade 10 donne par Yntema (1968) pour
Chelydra
serpentina:
embryons recti-
lignes, de 6-7 mm de longueur; ceil non pigmente; placode olfactive deprimee
en une cavite; bourgeon maxillaire superieur tres court; bourgeon allantoidien
non apparent exterieurement. Le developpement des embryons des stades 4 et 5
est comparable a celui du stade 11 d'Yntema: embryons a legere courbure
cervicale, de 7 mm de longueur; ceil pigmente; bourgeon maxillaire s'etendant
jusqu'au bord de l'oeil; bourgeon allantoidien non apparent exterieurement.
Les radioelements utilises sont la [3H]methyl thymidine (activites specifiques:
18 ou 25 Ci/mM), [5-3H]uridine (activites specifiques: 20 ou 6 Ci/mM), [4,5-3H]-
L-leucine (activite specifique: 15 Ci/mM), a la dose de 5 a
10
/*Ci environ par
oeuf,
pour chacun des radioelements. L'injection a ete faite en une seule fois,
pres du bord de l'aire vasculaire que Ton repere facilement par transparence a
travers la coquille en mirant
l'ceuf;
la quantite de liquide injectee est de 1/100 a
1/50 ml par
oeuf.
Les embryons traites a la thymidine tritiee ont ete sacrifies 4 h
apres l'injection (temperature d'incubation: 30 °C); ceux traites a la leucine

Autoradiographie
du membre d'embryons de
Cheloniens
587
tritiee, 2h apres l'injection (temperature d'incubation 25 °C); avec l'uridine
tritiee, la fixation des embryons a ete faite le plus souvent 2 h apres l'injection
(temperature d'incubation: 25 °C), pour quelques uns 4h apres (temperature
d'incubation 30
°C).
Le melange de Bouin a servi de fixateur. Les coupes, en serie
a
5
fim, ont ete trempees dans l'emulsion K
5
(Ilford) diluee de moitie et l'exposi-
tion a dure 12 a
14
jours. Apres developpement au Microdol X, une coloration
de fond a ete faite a l'hemalun-eosine. L'indice de marquage est obtenu en fai-
sant le rapport du nombre de noyaux marques a la thymidine tritiee au nombre
total de noyaux. Les comptages ont ete effectues, en general, sur la moitie des
coupes c'est-a-dire tous les 10/tm. L'indice de marquage est la moyenne des
comptages effectues a chaque niveau somitique. L'intensite du marquage a
l'uridine et a la leucine tritiee est appreciee par le nombre moyen de grains
d'argent par noyau (uridine) ou par cellule (leucine).
RESULTATS
Stade 1 {embryons de 21 a
23
paires de somites)
Du somite S6 au somite Slo (Sx designant le ler somite post-otique) et dan le
sens cranio-caudal, chaque somite envoie un prolongement ventral qui passe
entre l'epiblaste et le rein embryonnaire et vient au contact dufeuillet mesoder-
mique somatopleural a sa base. L'extremite du prolongement ventral du somite
touche Tangle laterodorsal de la cavite coelomique puis, s'en detachant et se
dirigeant vers l'epiblaste, il penetre a l'interieur du feuillet mesodermique soma-
topleural legerement epaissi, en poussant les cellules somatopleurales devant
lui (Fig. 1A); chez Tembryon d'Emys, cette extremite du prolongement somi-
tique est une vesicule dilatee qui souleve l'epiblaste localement avant de penetrer
dans le feuillet mesodermique somatopleural; elle est bordee de plusieurs couches
de cellules du cote de la future ebauche du membre, d'une seule couche de
cellules sur la face opposee; c'est une partie paraissant indifferenciee alors que,
a ce stade, la partie dorsale du somite est differenciee en dermatome et myotome.
Aux extremites des prolongements des somites S6 a S9, des cellules somitiques
se dissocient sans se separer encore du prolongement.
Le feuillet mesodermique somatopleural n'est epaissi qu'a la hauteur des
somites S6 a Slo qui sont seuls, a ce stade, a former des prolongements ventraux.
Ceci est vrai aux stades 22-23 paires de somites. Au stade 21 paires de somites,
cette description n'est valable que pour les somites S6 a S9 et l'epaississement de
la somatopleure ne se trouve qu'a ce niveau. Cet epaississement represente la
partie craniale du futur territoire de l'ebauche du membre; plus caudalement,
le feuillet mesodermique somatopleural, est reste constitue par un mince feuillet
de 2-3 couches de cellules.
Quelques differences regionales apparaissent a la fin de ce stade 1: au niveau
S6-S7, des cellules somatopleurales se sont tassees sous l'epiblaste; elles se
sont detachees du feuillet basal du mesoderme somatopleural; sous elles, le

588
J.
VASSE
D

Autoradiographie du membre d'embryons de Cheloniens 589
mesoderme somatopleural est plus lache. Ces phenomenes ne font que
commencer aux niveaux posterieurs. D'autre part la densite cellulaire est plus
faible dans la region S6-S7 que plus caudalement.
Marquage par la thymidine tritiee (Fig.
1
A)
D'une facon generate, l'extremite des prolongements soraitiques et le feuillet
mesodermique somatopleural sont beaucoup plus actifs (indices de marquage:
0,4 a 0,6) que l'epiblaste du territoire de la future ebauche du membre (indice
de marquage: 0,2 a 0,3). Les chiffres les plus eleves pour les indices de marquage
sont releves au niveau des somites S})-S10; le nombre limite d'embryons a ce stade
ne permet pas de savoir si cette difference est significative. Dans le feuillet
mesodermique somatopleural, sur un meme niveau transversal, on observe les
noyaux marques aussi bien dans le feuillet basal que juste au dessous de
l'epiblaste ou entre l'epiblaste et le feuillet basal du mesoderme somatopleural
(memes localisations que pour les mitoses).
D'autre part, on n'observe pas de difference significative, en ce qui concerne
les indices de marquage, entre l'epiblaste de la future ebauche de membre et
le reste de l'epiblaste embryonnaire, aux niveaux S6 a S10.
Marquage par V uridine tritiee
Le marquage, dans le cas d'une fixation de l'embryon deux heures apres
l'injection, est localise au niveau du noyau de la cellule marquee; pratiquement
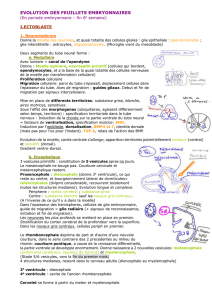
FIGURE 1
(A) Coupe transversale, passant par le 6eme somite (S6), d'un embryon d'Emys
de 22 paires de somites. Thymidine tritiee: l'extremite du somite (ex.S.) penetre
dans le mesoderme somatopleural
(m.s.).
Le feuillet mesodermique somatopleural
(m.s.) est nettement epaissi, bien marque. A l'extremite du somite, des cellules se
dissocient sans se separer encore du somite. ep.: epiblaste; r.\ rein embryonnaire.
(B) Coupe transversale, passant par
le
9eme somite
(Sd),
d'un embryon d'Emys de 26
paires de somites. Thymidine tritiee: trois zones sont visibles dans le mesoblaste: le
mesenchyme lache (m.l.) peu marque; le feuillet basal du mesoderme somato-
pleural (b.m.s.) et le mesoblaste basophile (m.b.) bien marques. Des cellules somi-
tiques (c.S.) se detachent de l'extremite du prolongement du somite (ex.S.).
(C) Coupe transversale, passant par le lOeme somite, d'un embryon de Testudo
de 34 paires de somites. Uridine tritiee; fixation 4h apres l'injection. Les assises
internes de la crete apicale
(c.a.)
ne presentent plus qu'un tres leger marquage malgre
la duree de Texposition. ep.: epiblaste.
(D) Coupe transversale, passant par le 8eme somite, d'un embryon d'Emys de 30
paires de somites. Thymidine tritiee. Dans l'epiblaste
(ep.),
faible marquage des
assises internes de la crete (c.a.: crete apicale). Dans le mesoblaste, nombreuses
cellules marquees dans le feuillet basal du mesoderme somatopleural
(b.m.s.),
pres
de Textremite du prolongement somitique (ex.S.).
(E) Coupe transversale, passant par le 9eme somite, d'un embryon d'Emys de 31
paires de somites. Uridine
tritiee.
La crete apicale
(c.a.)
est nettement moins marquee
que le reste de l'epiblaste
(ep.d.:
epiblaste dorsal: ep.v.: epiblaste ventral).
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%