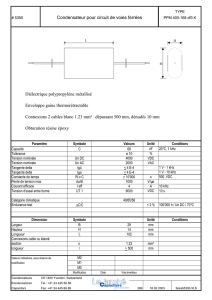
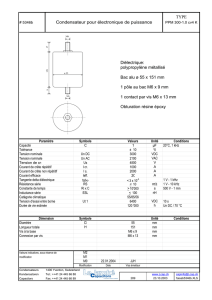
Le système auditif thalamo-cortical : anatomie, propriétés

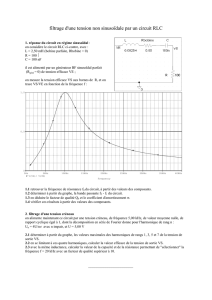
Troisième Partie:
dynamique du système thalamo-cortical,
influence des systèmes neuromodulateurs.

Les deux Systèmes Thalamo-Corticaux...
GABA
Glutamate
NBM
PPT
LDT
LC
Raphe
HT
Glu GABA
Sensory Cortex
Sensory
Reticular
Nucleus
Sensory
Thalamus
Sensory
Afferences
Vision
Somatosensory
Auditory
Sensory Cortex
Sensory Thalamus
Sensory
Reticular
Nucleus
MonoAminergic
Systems Cholinergic
System
NE 5HT
ACh HA

Les études in vitro indiquent que neuromodulateurs changent le mode de
décharge des neurones dans tout le réseau thalamo-cortical (McCormick 1992)
Neurone Thalamique
(McCormick & Prince 1988)
Neuromodulateurs
Neurone Cortical (Wang & McCormick 1993)
Neurone du Noyau Réticulaire Thalamique
(Pinault & Deschënes 1992)

Les effets des systèmes neuromodulateurs in vivo:
Effet de l’Acétylcholine dans le cortex auditif
30 40 50 60 70 80 90
Intensité (dB)
30 40 50 60 70 80 90
Intensité (dB)
Fonction Intensité Contrôle
Avec application d' ACh
Contrôle
ACh
Champ Récepteur Contrôle
Avec application d' ACh
Expansion du champ récepteur
(McKenna et al 1989) Diminution du seuil Acoustique
Metherate et al. (1990)

La Noradrénaline diminue les réponses évoquées dans le cortex auditif: elle
provoque un rétrécisemment des Chanps Récepteurs et une augmentation du
seuil acoustique (Manunta and Edeline, 1997, 1999)
.
10
8
6
4
2
Spikes / Bin
Control
NA 10nA
10
8
6
4
2
Spikes / Bin
8075706560555045403020
8075706560555045403020
Rétrécissement du champ récepteur Augmentation du seuil Acoustique
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
1
/
59
100%