Transgénèse

La transgenèse chez la souris
Souris transgénique classique: intégration aléatoire du transgène
dans le génome hôte = transgenèse additive
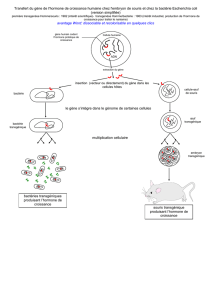
Souris transgéniques par recombinaison homologue: intégration ciblée
du transgène dans le génome hôte
Partie 1
Partie 2
Souris transgéniques générées par chimères d’agrégation
Partie 3

Qu’est-ce que la transgenèse?
Animal transgénique: animal dont le génome a été modifié
-soit par l’addition de nouvelles séquences d’ADN dans son génome
avec une intégration aléatoire
-soit par la modification de séquences déjà présentes dans son
génome (knock-out, knock-in) par recombinaison homologue
Ces modifications doivent être transmissibles par la lignée germinale d’une
génération à l’autre.

Pourquoi la transgenèse?
Permet d’étudier la fonction des gènes
-développement embryonnaire
-organogenèse
-pathologies humaines

Souris transgénique classique ou additive: intégration aléatoire du transgène
dans le génome hôte
Principes
Applications sur-expression d’un gène
expression ectopique d’un gène
analyse des régions régulatrices d’expression des gènes
« rescues »
Système inductible d’expression du transgène (tet-ON, tet-OFF)
Système conditionnel d’expression du transgène (CRE-loxP)
Approches siRNA (ou shRNA)
Partie 1

Souris transgénique classique: intégration aléatoire du transgène
dans le génome hôte
1-PRINCIPES
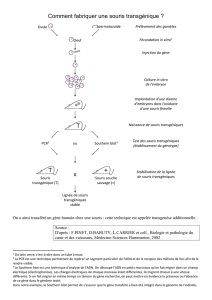
La transgenèse a pour but:
-d’introduire de manière stable (transmissible) un ou plusieurs gènes
exogènes dans le génome de l’animal hôte
L’introduction de l’ADN s’effectue:
- par micro-injection dans l’embryon au stade 1 cellule
partie 1
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
1
/
69
100%