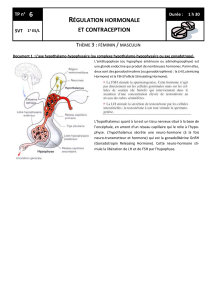
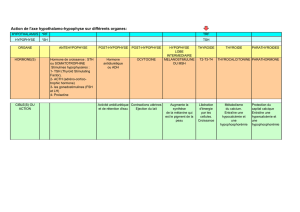
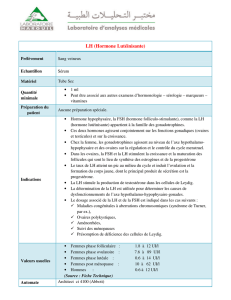
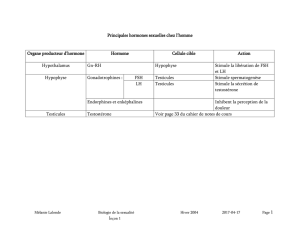
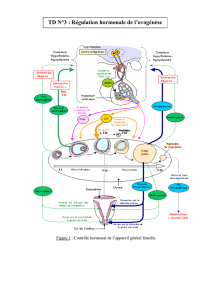
Régulation différentielle de l`expression des gonadotropines (LH et

THESE DE DOCTORAT DE
L’UNIVERSITE PIERRE ET MARIE CURIE
Spécialité : Physiologie de la Reproduction
Présentée par
Salima AROUA
Pour obtenir le grade de
DOCTEUR DE L’UNIVERSITE PIERRE ET MARIE CURIE
Régulation différentielle de l’expression des gonadotropines (LH
et FSH) chez l’anguille européenne, Anguilla anguilla
Soutenue le 9 juillet 2008
JURY
- Mme Le Gac Florence, DR INRA, Rennes Rapporteur
- Mr Kah Olivier, DR CNRS, Univ. Rennes Rapporteur
- Mr Boeuf Gilles, PR UPMC, Banyuls Examinateur
- Mr Counis Raymond, DR CNRS, Paris Examinateur
- Mr Elie Pierre, DR Cemagref, Bordeaux Examinateur
- Mme Dufour Sylvie, DR CNRS, MNHN Directeur de thèse
tel-00811792, version 1 - 11 Apr 2013

REMERCIEMENTS
Dans un premier temps, je voudrais remercier les membres du jury, Madame Florence Le Gac
et Monsieur Olivier Kah qui ont accepté « d’endosser les rôles de rapporteurs », ainsi qu’à
Messieurs Gilles Boeuf, Pierre Elie et Raymond Counis qui m’ont fait l’honneur d’accepter
de participer à mon jury de thèse et d’évaluer mon travail.
J’adresse mes plus sincères remerciements à Madame Sylvie Dufour qui m’a formée,
coachée, supportée, aidée et motivée au cours de toutes ces années passées au sein de son
laboratoire. Ce fut un réel plaisir, j’y ai appris énormément sur le plan scientifique comme sur
le plan humain.
Un grand merci également à l’ensemble de l’équipe qui m’a accueillie en son sein : à Nadine
qui m’a formée à la culture cellulaire et qui dans les derniers mois a été un de mes nombreux
coachs ; à Karine pour son aide et son soutien au cours des périodes de rédaction et en
dehors ; à Bernadette et Sylvie pour les travaux d’histologie et les dosages hormonaux, pour
bonne humeur quotidienne (ou presque) et pour les nombreuses discussions et fous rires
partagés ; et à Marie-Emilie avec qui j’ai partagé le « bateau de la thèse ». Merci, merci,
merci.
Je voudrais également exprimer ma reconnaissance à Madame Monika Schmitz qui m’a
initiée à la biologie moléculaire, il y a quelques années de cela et qui, tout au long de mon
parcours, m’a procuré de judicieux conseils. J’espère pouvoir en profiter encore longtemps.
Merci également à Monsieur Finn-Arne Weltzien qui a su, avec douceur et sagesse, me faire
pousser les portes de la PCR quantitative en temps réel…. Et qui, par la suite, m’a si
gentiment reçu au « pays des Vikings ».
Au cours de ces années, j’ai pu rencontrer une multitude de personnes avec lesquelles j’ai pu
échanger et m’enrichir tant d’un point de vue scientifique, que d’un point de vue humain.
Donc merci à l’équipe des Céphalos : Laure, Aude, Sébastien, Yann, Sandra et tous les gens
de passage pour ces petits-déjeuners studieux qui m’ont appris et m’apprennent encore, au fur
et à mesure, à améliorer mes discours au public.
Merci aux copains thésards ou jeunes docteurs: Gersende, Cécile, Hélène (la
« Cemagrefienne »), Anne-Gaëlle, Eric, Patrick (alias « speedy gonzalez »), Marie-Emilie,
Jean-Baptiste, Stéphanie, Sandra et tous les autres qui, partageant les mêmes angoisses,
épreuves, difficultés, mais aussi motivations et joies, m’ont permis d’avancer ; et aux
« autres » docteurs : Catherine, Miskal, Marthe,… qui me montrent que tout est possible.
Merci à tous les habitants (ou ayant habité) du 7 rue Cuvier. Merci pour les pauses-café et les
déjeuners dans la ménagerie, merci pour la bonne humeur quotidienne, merci pour le soutien ;
bref, je suis vraiment heureuse d’être passée par là.
Merci aux amis et amies qui m’ont soutenue et motivée pendant tout ce temps, m’écoutant
patiemment leur parler du cycle de vie de l’anguille, de mes travaux en cours ou encore de
mes chers élèves ; ça n’a pas dû être facile, je vous félicite donc pour cela et vous remercie du
fond du coeur….
tel-00811792, version 1 - 11 Apr 2013

Et bien sûr, un grand grand merci à ma famille. Mes sœurs, Saida, Karima et Lila, mes frère et
beau-frère, Salim et Jean-Marc, mes parents Mohand et Chala, et les nouveaux arrivants
Salma et Yanis qui m’ont porté et ont cru en moi tout au long de mon parcours.
Enfin, j’aimerais dédier ce travail à tous les miens et plus spécialement à mon grand père
Abdellah Aroua qui m’a appris que la patience et la sagesse étaient les plus sains objectifs à
atteindre.
tel-00811792, version 1 - 11 Apr 2013

Liste des publications et communications réalisées pendant la formation doctorale
Articles publiés:
Schmitz M, Aroua S, Vidal B, Le Belle N, Elie P, Dufour S, 2005. Differential regulation of
luteinizing hormone and follicle-stimulating hormone expression during ovarian development
and under sexual steroid feedback in the European eel. Neuroendocrinology 81: 107-119.
Aroua S, Schmitz M, Baloche S, Vidal B, Rousseau K, Dufour S, 2005. Endocrine evidence that
silvering, a secondary metamorphosis in the eel, is a pubertal rather than a metamorphic event.
Neuroendocrinology 82: 221-232.
Sbaihi M, Kacem A, Aroua S, Baloche S, Rousseau K, Lopez E, Meunier F, Dufour S, 2007. Thyroid
hormone-induced demineralisation of the vertebral skeleton of the eel, Anguilla anguilla. Gen
Comp Endocrinol. 151(1): 98-107.
Aroua S, Weltzien F-A, Le Belle N, Dufour S, 2007. Development of real-time RT-PCR assays for
eel gonadotropins and their application to the comparison of in vivo and in vitro effects of sex
steroids. Gen Comp Endocrinol. 153(1-3): 333-343.
Article en préparation:
Aroua S, Baloche S, Le Belle N, Rousseau N, Dufour S. Activin differentially regulates luteinizing
hormone and follicle-stimulating hormone expression by eel pituitary cells.
Chapitres de livre:
Rousseau K, Aroua S, Schmitz M, Elie P, Dufour S. Silvering: metamorphosis or puberty ? In
‘Migration and Reproduction of the European eel’ (G. van den Thillart, S. Dufour, C. Rankin,
eds), Springer Publisher (sous presse).
Aroua S, Rousseau K, Schmitz M, Chang CF, Dufour S. Positive and negative regulations of
endogenous brain gonadoliberins and pituitary gonadotropins during eel artificial maturation. In
‘Migration and Reproduction of the European eel’ (G. van den Thillart, S. Dufour, C. Rankin,
eds), Springer Publisher (sous presse).
Communication orale:
Aroua S, Schmitz M, Weltzien F-A, Baloche S, Le Belle N, Rousseau K, Dufour S, 2006. Differential
expression of LHbeta and FSHbeta in European eel, Anguilla anguilla. Proceedings of the 23rd
Conference of European Comparative Endocrinologists, Manchester, England. Abstract book p.
64, oral communication (O79).
Communications affichées:
Aroua S, Sbaihi M, Huang Y-S, Le Belle N, Dufour S, 2007. Synergistic action of androgens and
cortisol on gonadotropin regulation in European eel, Anguilla anguilla. Proceedings of the 8th
International Symposium on Reproductive Physiology of Fish in Saint Malo, France. Abstract
book p. 118, poster communication (P27).
Aroua S, Le Belle N, Schmitz M, Dufour S, 2007. In vitro regulation by activinB and IGFI of
gonadotropin expression in European eel, Anguilla anguilla. Proceedings of the 8th
International Symposium on Reproductive Physiology of Fish in Saint Malo, France. Abstract
book p. 119, poster communication (P28).
tel-00811792, version 1 - 11 Apr 2013

Principales abréviations
11-KT: 11-kétotestostérone
CA: catécholamine
DA: dopamine
DHP: 17!,20"-dihydroxy-4-pregen-3-one
DHT : dihydrotestostérone
E2 : oestradiol
EIA - ELISA: dosage immuno-enzymatique
F : Cortisol
FSH : hormone folliculo-stimulante (follicle-stimulating hormone)
FSH" : sous-unité " de la FSH
GH : hormone de croissance (growth hormone)
GnRH : gonadolibérine (gonadotropin-releasing hormone)
GP : hormone glycoprotéique
GP! : sous-unité ! des hormones glycoprotéiques
hCG : gonadotropine chorionique humaine
IGF : facteur de croissance de type insuline (insulin-like growth factor)
IO : index oculaire
LH : hormone lutéinisante (luteinizing hormone)
LH" : sous-unité ! de la LH
HPLC : chromatographie liquide haute pression
NA : noradrénaline
NPOav : noyau préoptique antéroventral
POA : aire préoptique
PRL : prolactine
qrt RT-PCR: PCR quantitative en temps réel
RIA: dosage radio-immunologique
RGS - GSI : rapport gonadosomatique (Poids des gonades / Poids total du corps)
RTDS – DSI : rapport tractus digestif-somatique (Poids du tractus digetif/ Poids total du corps)
T : testostérone
T3 : triiodothyronine
T4 : thyroxine
TSH : hormone thyréostimulante (thyroid-stimulating hormone)
TH : tyrosine hydroxylase
Vtg : vitellogénine
tel-00811792, version 1 - 11 Apr 2013
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
1
/
234
100%