PDF - 782.9 ko

La cellule eucaryote, unité structurale et fonctionnelle- 1
F. Brondex et E. Paitel
PARTIE 2. LA CELLULE DU PARENCHYME PALISSADIQUE
Introduction
Une plante est aussi un organisme pluricellulaire. Elle possède aussi des organes spécialisés : les racines assurent
l’ancrage et l’absorption de molécules du sol, les feuilles permettent les échanges gazeux et la réalisation de la
photosynthèse. Il existe alors des cellules spécialisées : quelles sont les particularités des cellules du parenchyme
palissadique, catégorie de cellules qu’on trouve dans les feuilles ? Quelles sont leurs caractéristiques
structurales ? Leur fonction est la réalisation de la photosynthèse. Rappel : en présence de lumière, le CO2 et l’eau
réagissent et sont convertis en molécule organique et O2. Quelles sont les caractéristiques fonctionnelles, quels
sont les caractères en relation avec la réalisation de la photosynthèse ?
Quels sont les différences et les points communs entre cellule animale et végétale ?
I. LA CELLULE DU PARENCHYME PALISSADIQUE DANS LA FEUILLE
A. Organisation d’une feuille
On peut distinguer dans la feuille :
- deux épidermes, un épiderme est constitué d’une seule couche de cellules jointives. Les cellules
épidermiques sont recouvertes d’une cuticule de nature lipidique. Cette cuticule a des propriétés
d’imperméabilité : elle limite l’évaporation de l’eau à la surface de la feuille. Il n’y a pas d’échanges gazeux
comme O2 ou CO2. Mais elle protège aussi la feuille contre l’entrée des micro-organismes…. Au niveau de
l’épiderme inférieur, on trouve des stomates. Les stomates sont constitués de deux cellules
(chlorophylliennes) en forme de haricot. Elles peuvent s’écarter plus ou moins laissant un petit espace :
l’ostiole. Les stomates sont les lieux d’échanges de gaz pour la feuille : l’eau sous forme de vapeur sort de la
feuille, les principaux échanges de gaz concernent le O2 ou CO2.
- Entre les deux épidermes, on trouve le mésophylle, constitué de :
Un parenchyme lacuneux : les cellules sont chlorophylliennes (elles contiennent des chloroplastes).
Elles ne sont pas totalement jointives. Entre les cellules, les gaz peuvent circuler.
Des faisceaux conducteurs correspondent aux nervures de la feuille. Les faisceaux conducteurs sont
formés des vaisseaux de transport de la sève. Les vaisseaux du xylème transportent la sève brute, des
racines vers les feuilles. Le phloème distribue la sève élaborée des feuilles vers les organes
consommateurs.
Le parenchyme palissadique (en position supérieure) est aussi constitué de cellules chlorophylliennes,
mais jointives, très géométriques.
On remarque donc des relations structure fonction à l’échelle de la feuille : limbe aplati comme surface de
réception de la lumière, les cellules du parenchyme palissadique sont donc situées dans une feuille, où les gaz
circulent facilement (grâce aux stomates et aux lacunes du mésophylle), où l’eau nécessaire à la photosynthèse
est apportée par les vaisseaux du xylème.
Ainsi, une cellule du parenchyme palissadique n’est pas isolée mais intégrée dans un organe et un organisme ce
qui implique des échanges (eau, CO2, O2…)
B. Les limites cellulaires : paroi et membrane plasmique
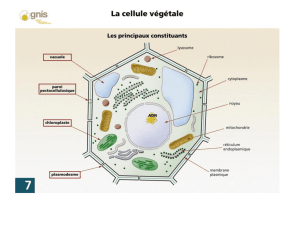
La cellule végétale est délimitée par une membrane plasmique, très semblable à celle d’une cellule animale. Elle
est constituée d’une bicouche de lipides et de protéines (propriétés des mb détaillées ultérieurement). Elle joue
aussi un rôle de barrière sélective. Mais la mb des cellules végétales est en général non observable car elle est
accolée à la paroi de la cellule.
Remarque : on parle aussi de plasmalemme.
La paroi est propre aux cellules végétales, elle est de nature pecto-cellulosique : cellulose et pectines qui sont des
dérivés glucidiques. Deux cellules adjacentes mettent en commun les parois qu’elles ont fabriquées.
La paroi forme un cadre rigide autour de la cellule, elle assure la fonction de squelette extracellulaire :
- elle impose sa forme à la cellule
- elle donne rigidité à la cellule, c’est-à-dire qu’elle limite sa déformation (cf. cellule turgescente, cellule en
plasmolyse)

La cellule eucaryote, unité structurale et fonctionnelle- 2
F. Brondex et E. Paitel
Par ailleurs, elle n’empêche pas complètement la croissance de la cellule (qui peut être vue comme un
changement de forme). Dans une jeune cellule, la paroi est suffisamment déformable : sous l’effet de la pression
de turgescence de la vacuole, la paroi se déforme et la cellule peut grandir.
Bilan : la paroi joue le rôle de squelette de la cellule végétale.
Ne pas oublier de placer la membrane
Les plasmodesmes sont des structures qui permettent la connexion des cytoplasmes de 2 cellules adjacentes. La
paroi est interrompue au niveau d’un plasmodesme (ponctuation). La ponctuation est tubulaire. La mb est
accolée à la paroi, y compris au niveau de la ponctuation. Le RE traverse la ponctuation, on parle de desmotubule.
Entre la mb plasmique et le desmotubule, il y a un ensemble de protéine en anneau. Au niveau d’un
plasmodesme, des petites molécules comme des ose ou des acides aminés peuvent traverser d’une cellule à
l’autre. Les ions monovalents peuvent aussi traverser le plasmodesme. Par contre, les grosses molécules sont trop
encombrantes et ne passent pas. Les influx électriques enfin (dépolarisation ou hyperpolarisation) peuvent passer
d’une cellule à l’autre via les plasmodesmes.
Cellule 1
Cellule 2
Cellule 3
méat
Lamelle moyenne
(pectines)
Paroi cellule
Dessiner les
cellules entières

La cellule eucaryote, unité structurale et fonctionnelle- 3
F. Brondex et E. Paitel
C. La cellule du parenchyme palissadique : une cellule eucaryote
1) Une vue d’ensemble
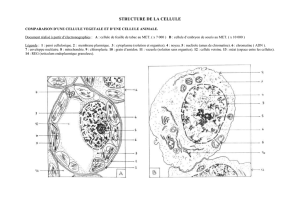
Ultrastructure de la cellule (à partir MET) : la taille est importante environ 100µm de long. On remarque des
particularités propres aux cellules eucaryotes. En effet, Le cytoplasme est compartimenté, il contient des
organites communs aux cellules animales et végétales :
- un noyau, une enveloppe (double mb percée de pores) délimitant un nucléoplasme. L’ADN est contenu
dans le noyau et apparaît sous forme de chromatine. On distingue une zone plus dense aux électrons, le
nucléole.
- Le REG, REL et l’appareil de Golgi. On remarque un moindre développement par rapport à la cellule
acineuse pancréatique, la synthèse protéique existe mais elle n’est pas la fonction principale.
- Des mitochondries sont présentes : elles sont aussi le siège de la respiration cellulaire et de la production
d’ATP.
Des organites spécifiques des cellules végétales sont présents : vacuole, chloroplastes…
Les cellules végétales possèdent aussi un cytosquelette avec les mêmes constituants moléculaires que les cellules
animales (autre caractère d’Eucaryotes)
Le cytoplasme est réduit et superficiel, en rapport avec la présence d’une vacuole. Par conséquents, les
différentes zones cellulaires sont éloignées (en particulier en rapport avec la grande taille de la cellule) : la
diffusion classique des molécules est peu ne permet pas une distribution homogène.
Or, il existe un courant cytoplasmique orienté, entraînant les molécules, les organites (cf chloroplastes). Ce
courant se met en place autour de la vacuole : c’est la cyclose. Il repose sur une structuration du cytoplasme, on
distingue : l’ectoplasme (plus externe et plus visqueux) et l’endoplasme (plus interne et moins visqueux).
La mise en mouvement de l’ectoplasme par rapport à l’endoplasme nécessite de l’énergie sous forme d’ATP. A
l’interface entre les deux zones, on trouve des protéines du cytosquelette : actine côté ectoplasme, myosine côté
endoplasme. L’interaction entre les deux permet le mouvement.
Mb plasmique
vacuole
endoplasme
ectoplasme
paroi
Structure du cytoplasme
paroi
mb
RE

La cellule eucaryote, unité structurale et fonctionnelle- 4
F. Brondex et E. Paitel
2) Les péroxysomes, des organites présents dans toutes les cellules eucaryotes
Les péroxysomes sont des organites à simple mb, d’environ 1µm de diamètre. Les péroxysomes sont présents
dans toutes les cellules eucaryotes, végétales ou animales. Ils sont impliqués dans le métabolisme de l’oxygène
dans les cellules et des réactions de détoxification. Certaines molécules produites par le métabolisme peuvent
être toxiques, elles sont alors modifiées par des réactions chimiques particulières. Ces réactions impliquent le
dioxygène (O2). Le dioxygène est utilisé par les mitochondries (respiration) et produit par la photosynthèse dans
les chloroplastes. Le dioxygène est un fort oxydant et peut lui aussi sous des formes dérivées devenir toxique pour
la cellule.
Une molécule RH2 est à dégrader grâce à des enzymes spécifiques :
RH2 + O2 R + H2O2 (peroxyde d’hydrogène lui-même potentiellement toxique)
La catalase utilise le peroxyde d’hydrogène pour oxyder d’autres molécules potentiellement toxique :
H2O2 + R’H2 R’ + 2H2O
Eventuellement, le peroxyde d’hydrogène s’accumule, la catalase peut catalyser la réaction suivante :
2H2O2 2H2O + 2O2
Les enzymes sont parfois tellement concentrées qu’elles forment des structures cristallines.
Remarque : chez les animaux, on trouve ces réactions de détoxification, par exemple, les péroxysomes des
cellules de foie dégradent des molécules comme l’éthanol selon la même voie.
Chez les végétaux, le péroxysome est aussi le lieu de la βoxydation : une série de réaction qui dégradent les acides
gras (lipides), ce qui permet une importante récupération d’énergie (en coopération avec les mitochondries).
Enfin, les réactions de la photorespiration (non détaillées ici) sont réalisées par le péroxysome, le chloroplaste et
la mitochondrie en coopération.
II. LA CELLULE DU PARENCHYME PALISSADIQUE POSSEDE DES ORGANITES SPECIFIQUES AUX CELLULES
VEGETALES
A. Les chloroplastes, siège de la photosynthèse et de l’autotrophie au carbone
Les plastes sont des organites à double mb. Il existe différentes catégories de plastes à rôles différents, dans la
cellule du parenchyme palissadique, on trouve des chloroplastes, spécialisés dans la photosynthèse (cf ce
chapitre).
Chez l’épinard, la taille moyenne d’un chloroplaste est 5 par 2µm. L’observation au MET montre l’ultrastructure
d’un plaste :
La mb est double, elles délimitent un espace intermembranaire de composition assez proche du cytosol car la mb
externe est assez perméable.
Dans le chloroplaste, on trouve un réseau de mb internes orientées parallèlement : les thylakoïdes. Les mb
thylakoïdiennes délimitent un espace clos et continu : le lumen. Les mb thylakoïdiennes sont disjointes de la mb
interne. Les mb thylakoïdiennes contiennent les complexes pigmentaires impliqués dans la photosynthèse comme
la chlorophylle (verte) : les pigments contenus dans les thylakoïdes permettent d’absorber la lumière. On trouve
aussi des ATP synthases dans les mb thylakoïdiennes. On distingue des thylakoïdes granaires (empilés) et
intergranaires. La surface « plissée » des thylakoïdes développe une grande surface de réception de la lumière :
exemple de relation structure fonction.
Un troisième compartiment est délimité : le stroma, il contient un certain nombre de molécules particulières.
- Le stroma contient de l’ADN (un chromosome circulaire), des ribosomes, comme la mitochondrie, le
chloroplaste est dit semi autonome, il contient sa propre information génétique. La présence d’ARNr, ARNm,
ARNt montre qu’il y a synthèse de protéines.
- Les grains d’amidon (un polymère de glucose) correspondent à un stockage temporaire des produits de la
photosynthèse, le jour en conditions d’intense éclairement. La nuit cet amidon est dégradé, les glucides sont
exportés.
- Il y aussi des gouttelettes de lipides et de caroténoïdes (pigments lipidiques) : il s’agit d’un stock de lipides qui
permet l’entretien des mb internes du chloroplaste.
- Enfin, on trouve des enzymes particulières comme la Rubisco, enzyme capitale de la photosynthèse. De façon
générale, on trouve des enzymes impliquées dans la photosynthèse ou la synthèse des acides aminés (2 fonctions
du chloroplaste)
Ainsi le chloroplaste est le siège de la photosynthèse (équation) permettant l’autotrophie pour le carbone de la
plante.

La cellule eucaryote, unité structurale et fonctionnelle- 5
F. Brondex et E. Paitel
On peut remarquer aussi que les molécules comme les enzymes sont ainsi plus concentrées dans le petit volume
du chloroplaste que dans le volume total de la cellule (effet compartimentation).
Mais il existe d’autres membres de la famille des plastes (on ne les trouve pas dans la cellule du parenchyme
palissadique)
Les amylopastes sont les organites de stockage de l’amidon à plus long terme comme dans les organes de
réserves (exemple Pomme de Terre). On retrouve la double membrane mais pas de thylakoïdes. Le stockage dans
un compartiment délimité par des mb permet un stockage sans modification de la pression osmotique du cytosol
(encore la compartimentation !)
Les chromoplastes contiennent dans leurs mb de nombreux pigments de type lipidiques, par exemple des
caroténoïdes de couleurs jaune ou orange.
Les plastes appartiennent à une même famille et peuvent dériver les uns des autres.
B. La vacuole, un organite aux fonctions diverses
La vacuole est un organite souvent volumineux, qui rejette le cytoplasme et les autres organites en périphérie.
Mais la taille de la vacuole dépend du type cellulaire. D’une façon un peu générale, les cellules peu différenciées
contiennent plusieurs petites vacuoles, alors que les cellules plus différenciées ont une seule grande vacuole.
La vacuole est délimitée par une simple mb, constituée de lipides et de protéines : le tonoplaste. Les propriétés
de cette mb sont similaires à celles de la mb plasmique, elle constitue une barrière à perméabilité sélective : elles
contient des transporteurs permettant les échanges de molécules avec le cytosol. La composition de la solution
de la vacuole est assez différente de celle du cytoplasme.
- La vacuole contient une solution aqueuse. L’essentiel de l’eau de la cellule est contenue dans la vacuole. Celle-ci
joue un rôle dans l’équilibre hydrominéral de la plante (contenu en eau et en ions). On a observé en TP la
turgescence et la plasmolyse d’une cellule selon la pression osmotique du milieu. L’eau exerce une pression sur la
paroi de la cellule, c’est la pression de turgescence. Lorsque la plante est bien hydratée, les cellules sont
turgescentes, la pression de turgescence est assez élevée contre la paroi. La pression de turgescence permet de
maintenir le port dressé de la plante (cf. pression dans un tuyau d’arrosage). En cas de stress hydrique, la vacuole
diminue de volume, la cellule est en plasmolyse.
Par ailleurs, lorsque la cellule est non différenciée, sa paroi est encore peu épaisse et elle est encore élastique.
Elastique : revient à sa forme d’origine si elle est déformée. Dans ces conditions, si la cellule est turgescente alors
la vacuole fait pression sur la paroi. Celle-ci se déforme et la cellule peut augmenter sa taille.
- Les molécules en solution : la composition du liquide vacuolaire est variable selon l’espèce, le type cellulaire, ou
l’état physiologique mais on peut retenir quelques données générales :
Les glucides, par exemple le malate ou le saccharose (produits indirects de la photosynthèse). On mesure en
nmol/mg de chlorophylle dans une feuille d’épinard.
Résultats : La quantité totale de saccharose ou de malate augmente au cours de la journée (à la lumière) puis
diminue à l’obscurité. La quantité de saccharose ou de malate augmente essentiellement dans la vacuole puis
elle diminue ensuite au retour à l’obscurité, elle reste à peu près constante et faible dans le cytosol et le stroma.
Interprétation : Les quantités augmentent car ce sont des produits de la photosynthèse. Ces molécules
s’accumulent dans la vacuole la journée : au fur et à mesure de la production, le malate et le saccharose sont
stockés dans la vacuole. Ce stockage est temporaire puisque à l’obscurité, les quantités diminuent à nouveau.
Les molécules sont exportées vers des lieux de stockage à plus long terme ou vers des organes consommateurs.
On peut remarquer que les molécules ne sont pas stockées dans le cytosol. En effet, le stockage dans la vacuole
permet de ne pas modifier la pression osmotique du cytosol. On voit encore une des conséquences de la
compartimentation.
Remarque : l’accumulation est particulièrement importante et plus durable chez la canne à sucre et la betterave,
qui sont exploitées pour le sucre alimentaire.
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%