Organisation laminaire du cortex strié

Vision: organisation anatomo-
fonctionnelle des voies centrales

1-Projection rétinofuge
•Correspond au prolongement des nerfs optiques;
•Rétinofuge, du latin fugere: fuir;
•Étude du système visuel central (SVC) -> observation du
trajet des voies rétinofuges, des yeux jusqu’au tronc
cérébral et le traitement effectué dans ces voies.


1-Projection rétinofuge
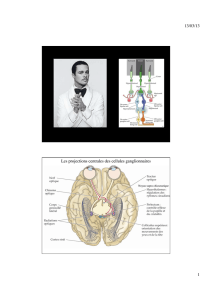
Nerf optique, chiasma optique et
tractus optique
•Axones des cellules ganglionnaires passent dans 3
structures avant de faire synapse dans tronc cérébral: nerf
optique, chiasma optique et tractus optique
•Nerfs optiques: quittent yeux à partir des disques
optiques, traversent les orbites et pénètrent dans le crâne;
•Chiasma optique: à la base du cerveau, en avant de
l’hypophyse; décussation partielle des axones provenant de
la rétine nasale;
•Tractus optique: sous la pie-mère, le long de la surface
latérale du diencéphale.

1-Projection rétinofuge
Hémichamps visuels droit et gauche
•Champ visuel: espace visible quand les deux yeux regardent droit
devant
•Hémichamp: champ visuel divisé par une ligne imaginaire passant en
son plein centre;
•La partie centrale des deux hémichamps visuels se forme sur les deux
rétines à la fois -> champ visuel binoculaire;
•Objets situés dans la partie binoculaire de l’hémichamp visuel gauche
sont reproduits à la fois sur la rétine nasale gauche et la rétine
temporale droite;
•Fibres nerveuses de la rétine nasale gauche passent du côté droit
(controlatérale) dans la décussation du chiasma optique;
•Fibres nerveuses de la rétine temporale droite poursuivent leur trajet de
façon ipsilatérale (du même côté);
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
1
/
57
100%