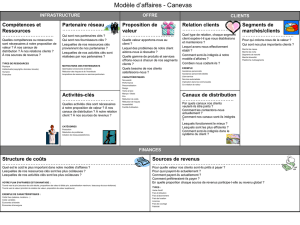
fiziologie cardiacă

Walther Bild, MD, PhD

Physiologie du myocarde
le métabolisme cardiaque
propriétés fondamentales du muscle
cardiaque
La révolution cardiaque
L’hémodynamique intracardiaque
Événements mécaniques de la révolution
cardiaque
Les manifestations acoustiques de la
révolution cardiaque
manifestations électriques de la
révolution cardiaque
Electrocardiographie
Le débit cardiaque
Le travail du cœur
Réglage de l'activité cardiaque
Physiologie de la circulation
artérielle
Notions de hémodynamique
particularités morphologiques et
fonctionnelles du système artériel
La pression artérielle
Le pouls artériel
Pouls volumétrique
Physiologie de la circulation
veineuse
particularités morphologiques et
fonctionnelles du système veineux
hémodynamique veineuse
la pression veineuse
temps de circulation
réglage de la circulation veineuse
La physiologie de la circulation
capillaire
particularités morphologiques et
fonctionnelles des capillaires sanguins
hémodynamique capillaire
échanges trans-capillaires
réglage de la circulation capillaire
Physiologie de la circulation
lymphatique
Morphologiques et particularités
fonctionnelles du système lymphatique
Le rôle de la circulation lymphatique


Les canaux ioniques dans le myocarde
•voltage-dépendants
•Na + (rapides)
•T Ca2 + (transitoire)
•L Ca2 + (longue durée)
•K +
•rectification interne
•rectification retardée
•efflux transitoire
•Canaux de K+ dépendantes de
récepteurs:
•Activées par le Ca2 +
•Activées par le Na+
•Sensibles à l'ATP
•activées par l'acétylcholine
•Activées par l'acide arachidonique
Le transport actif
au niveau du
cardiomyocyte
L’ATP-ase Na-K
Ca ATP-ase
L’Antiport Na/Ca

L’EXCITABILITÉ - FONCTION BATMOTROPE
L’AUTOMATISME - FONCTION CHRONOTROPE
LA CONDUCTION - FONCTION DROMOTROPE
LA CONTRACTILITE - FONCTION INOTROPE
LA TONICITE - FONCTION TONOTROPE
LA TROPHICITÉ - FONCTION TROPHOTROPE
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
1
/
29
100%