presentation au format ppt Fichier

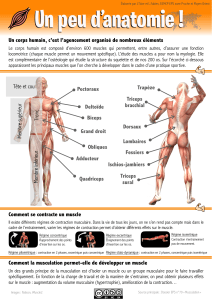
Le tissu musculaire des
Vertébrés
Structure et propriétés
Jean-Pierre GASC
UMR 7179
2013
SEP30

LES VERTÉBRÉS EFFECTUENT DES MOUVEMENTS
VARIÉS ET COMPLEXES DANS LEUR VIE QUOTIDIENNE
CES MOUVEMENTS REPOSENT SUR UN TISSU
PRODUCTEUR DE FORCES : LE MUSCLE SQUELETTIQUE
Photo A.R.DEVEZ Vidéo J.P.GASC

CHEZ TOUS LES VERTÉBRÉS, LES MUSCLES
REPRÉSENTENT UNE PART IMPORTANTE DE
LA MASSE DU CORPS.
Les félins rivalisent en record avec les saumons,
avec près de 60% de la masse
du corps dévolus aux muscles locomoteurs.

Cependant, la répartition de ces masses
varie beaucoup selon les adaptations
Antilocapra americana
Spalax erhenbergi
Course
Fouissage

La motilité propre à toute cellule
devient une spécialisation
dans la lignée des cellules du
muscle.
Les muscles sont les moteurs responsables
du déplacement et de la prise de nourriture
Leur structure repose sur un principe géométrique linéaire
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
1
/
51
100%