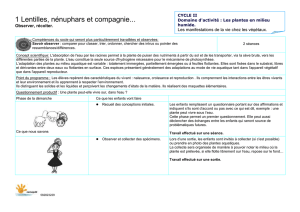
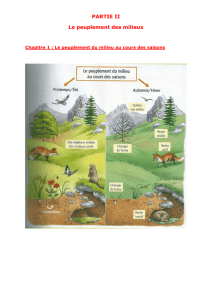
Quand les plantes se parlent

Édition 2014
Quand les plantes se parlent
Colloque scientifique
23 mai 2014 • Paris
Colloques scientifiques
et journées de conférences et d’échanges

16e colloque scientifi que - Paris, 23 mai 2014
QUAND LES PLANTES SE PARLENT
> Sommaire
Page 4 • La communication chez les plantes : un regard historique
Stéphane Tirard
Page 9 • Les outils de la communication végétale
Yvan Kraepiel
LA COMMUNICATION INTERNE
Page 14 • Les signaux d’un développement harmonieux
Arnould Savouré
Page 18 • Réactions aux stress environnementaux
Thierry Améglio
Page 25 • Le dialogue porte-greffe - greffon
Sarah Cookson
Page 30 • Impact de la nutrition azotée et de la variété du porte-greffe sur la
croissance végétative de la vigne.
Julien Lecourt
LA COMMUNICATION EXTERNE
Page 33 • L’exploitation des plantes par les plantes
Philippe Delavault
Page 38 • La symbiose, un échange gagnant-gagnant
Francis Martin
Page 41 • Les composés volatiles donnent l’alerte chez les plantes
Xavier Daire
Page 44 • Induction de résistances chez le blé lors d’une interaction com-
patible avec Blumeria graminis
Christine Tayeh
Page 51 • Insectes et plantes : je t’aime moi non plus
Frédéric Marion Poll
Page 54 • Conclusion
Yvan Kraepiel
Page 58 • Sélection de livres

4
La communication chez les plantes :
un regard historique
Stéphane Tirard, Centre François Viète d’Histoire des Sciences et de Techniques
Université de Nantes
Résumé
La communication chez les plantes renvoie à une
fonction évidente chez les animaux, mais beaucoup
plus problématique chez les végétaux. Le cadre de
notre réfl exion sera principalement celui de l’his-
toire des travaux sur la distinction entre les règnes
animal et végétal au XIXe siècle. L’idée de la «sen-
sibilité» des végétaux est en eff et à cette époque au
cœur de ce questionnement.
Nous analyserons donc les travaux d’un ensemble
de biologistes du XIXe siècle : Lamarck, Dutrochet,
Darwin, Bernard… afi n de révéler la complexité des
diff érences et analogies qu’ils établissent entres les
deux règnes, en mobilisant notamment la présence
ou l’absence de «sensibilité», tant par des constats
naturalistes que par des approches physiologiques.
Pour terminer, nous montrerons comment les
développements de la physiologie végétale durant
la deuxième partie du XIXe et le début du XXe siècle
ont révélé des spécifi cités fondamentales des végé-
taux sur lesquelles se fonde aujourd’hui la théma-
tique de ce colloque.
Introduction
Le présent colloque est intitulé : « Quand les plantes
se parlent ». Cette courte phrase révèle la relation
complexe qui nous lie aux végétaux. Mais elle nous
rappelle également la distinction établie de très longue
date entre ce que l’on appelait les deux règnes.
Cette phrase conduit à rouvrir le dictionnaire au mot
parler, si trivial que nous pourrions en oublier la signi-
fi cation. Il nous est confi rmé un sens premier corres-
pondant à l’usage de la voix chez l’homme : «Émettre
les sons articulés d’une langue maternelle ». Les plantes
semblent exclues de cette possibilité, tous les animaux
également d’ailleurs. C’est évidemment dans un sens
métaphorique que nous comprenons le titre de cette
journée. Il s’agirait d’étudier ici les mécanismes de
communication existant entre les plantes, en les assi-
milant à la parole. Ainsi, en se référant à cette fonction
de communication, ce titre compare les plantes aux
animaux, et notamment à l’homme.
Par vocation, le regard de l’historien des sciences se
porte sur le passé et donc nous reviendrons sur l’his-
toire de cette tension « diff érence / ressemblance »
entre les animaux et les végétaux, dans laquelle doit
être considérée cette métaphore sur « la parole » des
plantes. Nous commencerons par nous intéresser à
la limite végétal-animal telle qu’elle fut établie aux
XVIIIe et XIXe siècles. La présentation de quelques
étapes de la biologie, qui aux XIXe et XXe siècles, ont
montré l’unité du monde vivant à l’échelle cellulaire et
moléculaire, nous occupera dans un deuxième temps.
Enfi n, ceci nous permettra de saisir dans quelles
conditions il a été possible de montrer que la commu-
nication à l’intérieur de l’organisme végétal consiste
en des phénomènes moléculaires.
La limite végétal-animal
La distinction entre les règnes animal et végétal n’a
jamais manqué de poser un problème profond aux
observateurs, qu’ils soient naturalistes ou philo-
sophes. Je ne commencerai cette courte présentation
qu’au XVIIIe siècle, alors qu’il ne faudrait pas oublier
les auteurs qui, dès l’antiquité, ont souligné les diff é-
rences entre les deux règnes.
C’est Denis Diderot (1713-1784) lui-même qui signe
l’article intitulé Animal dans l’Encyclopédie. Il y étudie
notamment les diff érences avec le végétal :
« L’animal réunit toutes les puissances de
la nature ; les sources qui l’animent lui sont
propres et particulières ; il veut, il agit, il se
détermine, il opère, il communique avec ses
sens avec les objets les plus éloignés ; son indi-
vidu est un centre où tout se rapporte ; un point
où l’univers entier se réfl échit ; un monde en
raccourci.1»
C’est ainsi la relation aux autres êtres que Diderot
place au centre de sa défi nition. Le lecteur peut donc
s’attendre à la retrouver comme un critère de distinc-
tion entre les deux règnes dans la suite de son analyse.
Plus loin, Diderot s’appuie en eff et sur la propriété
d’irritabilité :
« Je ne crois pas que la fi bre végétale ait la faculté
d’être irritable. Si l’on observe dans beaucoup
de végétaux des phénomènes qui semblent
1 Denis Diderot, “Animal”, Encyclopédie ou dictionnaire raisonné des sciences et des arts et des métiers, Paris, Flammarion, 1986, p. 242.

5
indiquer une véritable irritabilité, comme les
mouvements singuliers des sensitives (mimosa
pudica, oxalis sensitiva, etc.) dans les articula-
tions de leurs rameaux ou de leurs feuilles, et
celui des étamines de l’épine-vinette (berberis
vulgaris), etc. Lorsqu’on touche ces parties ou
qu’on les ébranle d’une manière quelconque ; je
suis persuadé que ces mouvements sont dus à
une cause mécanique, et non à l’irritabilité des
parties.2 »
Le philosophe attribue ainsi à l’irritabilité un rôle cru-
cial dans la distinction entre les deux règnes et elle est
donc éliminée chez les végétaux.
Pour les observateurs naturalistes, le problème de la
limite animal-végétal s’est posé à maintes reprises aux
XVIIe et XVIIIe siècles, notamment dans le cadre de
la classifi cation. C’est ainsi que les zoophytes, par leur
apparence, semblent de part et d’autre de cette fron-
tière et leur dénomination entretint longtemps l’ambi-
guïté. En 1801, Jean-Baptiste Lamarck (1744-1829)
rappelait ce débat, qui a perduré, par un commentaire
tranché :
« La connaissance de ces animalcules et la
considération des masses3 ordinairement
rameuses et dendroïdes qui leur servent de
réceptacle et d’habitation, fi rent ensuite don-
ner à ces mêmes masses le nom de zoophytes,
qui veut dire animaux-plantes, comme si les
objets dont il s’agit participaient de la nature
de l’animal et de celle de la plante. On a même
prétendu, dans des ouvrages très modernes,
que les polypiers rameux croissaient par
intussusception4 , en sorte que le tronc et les
branches étaient de véritables végétations et
leurs auteurs ont donné le nom de fl eur-animal
au polype même qui habite chaque cellule de
ces polypiers. Mais cette opinion est une erreur
évidente. Il n’y a dans le polypier le plus rami-
fi é rien qui tienne de la nature d’un végétal ; si
l’on en excepte l’apparence ou la confi guration
extérieure. Tout y est animal ou production
animale. Chaque polype est un être vivant,
doué du mouvement volontaire et muni d’un
canal intestinal, aucun végétal connu n’off re
rien de semblable. »5
Qu’en est-il de la parole ? Elle n’est présente que chez
l’homme au terme d’une échelle de la complexité des
façons de communiquer.
« S’étant ainsi répandu presque partout, et
ayant pu se multiplier considérablement, ses
besoins s’accrurent progressivement par suite
de ses relations avec ses semblables, et se
trouvèrent infi niment diversifi és. Or, ceux des
animaux qui jouissent comme lui des facultés
d’intelligence, mais dans des degrés fort infé-
rieurs, n’ayant qu’un petit nombre de besoins
comparativement aux siens, n’ont aussi qu’un
très petit nombre d’idées ; et, pour communi-
quer entre eux, quelques signes leur suffi sent
entièrement. Il en est bien autrement à l’égard
de l’homme ; car ses besoins s’étant infi niment
accrus et diversifi és, et le forçant à multiplier
et à varier proportionnellement ses idées, il fut
obligé d’employer des moyens plus compliqués
pour communiquer sa pensée à ses semblables.
De simples signes ne lui suffi rent plus. Il lui
fallut non seulement varier les sons de sa voix,
mais en outre les articuler ; et selon le déve-
loppement particulier de l’état intellectuel de
chaque peuple, les sons articulés, destinés à
transmettre les idées, reçurent une complica-
tion plus ou moins grande. La faculté de for-
mer des sons articulés, qui, par convention,
expriment des idées, constitue donc celle de la
parole que l’homme seul a pu se procurer ; et la
nature des conventions admises, pour attribuer
à ces sons articulés des idées usuelles, constitue
aussi les diverses langues dont il fait usage. »6
Retenons donc que les débats naturalistes et philo-
sophiques du XVIIIe et du début du XIXe siècles ont
délimité les deux règnes. La distinction établie pou-
vait porter sur un très large ensemble de caractères, la
communication apparaissant alors comme l’apanage
des animaux et la parole comme celui de l’homme.
2 J.B. Lamarck, Mémoires de physique et d’histoire naturelle, établis sur les bases de raisonnement indépendantes de toute théorie ; avec
l’explication de nouvelles considérations sur la cause générale des dissolutions ; sur la matière de feu ; sur la couleur des corps ; sur la
formation des composés ; sur l’origine des minéraux, et sur l’organisation des corps vivants, lus à la première classe de l’Institut national
dans ses séances ordinaires, suivis de Discours prononcé à la Société Philomatique le 23 fl oréal an V, 1797, p. 12.
3 Masse : Groupe d’organismes
4 Intussusception : Pour les auteurs de l’époque, mode d’accroissement par pénétration des éléments nutritifs au sein des êtres organisés.
5 J.B. Lamarck, Système des Animaux sans vertèbres, ou Tableau général des classes, des ordres et des genres de ces animaux ; présentant
leurs caractères essentiels et leur distribution d’après les considérations de leurs rapports naturels et de leur organisation, et suivant
l’arrangement établi dans les galeries du Muséum d’Histoire naturelle, parmi leurs dépouilles conservées ; précédé du Discours d’ouver-
ture de l’an VIII de la République, Paris, Deterville, 1801, pp. 366-367.
6 J.-B. Lamarck, Système analytique des connaissances positives de l’homme, restreintes à celles qui proviennent directement ou indirec-
tement de l’observation, Paris, Chez l’Auteur et Belin 1820, pp. 151-152.

6
De la cellule à la molécule : une
nouvelle unité du vivant
Si nous venons de souligner que les végétaux sont défi -
nitivement distingués des animaux au début du XIXe
siècle, qu’en est-il de l’unité du monde vivant ? C’est
l’étude microscopique et moléculaire des organismes
qui la révèlera.
L’invention du microscope au XVII
e siècle n’a pas
entraîné automatiquement la découverte et la
conception d’une structure élémentaire microscopique
universelle du vivant. Les débats eff ectifs sur ce
problème ne datent en eff et que du premier tiers du
XIXe siècle. Ils concernent alors autant le végétal que
l’animal. Il est bien connu que la première théorie
cellulaire est due aux travaux complémentaires d’un
botaniste et d’un physiologiste. En eff et, le botaniste
allemand Mathias Schleiden (1804-1881) note que
les végétaux sont constitués de cellules structurées
autour d’un noyau, alors appelé le cytoblaste. Cette
idée est reprise par eodor Schwann (1810-1882) qui,
en 1839, l’étend aux animaux après de nombreuses
observations. Nous assistons donc ici à un phénomène
de généralisation d’un concept qui tend à montrer
l’unité du monde vivant.
La théorie cellulaire est révisée dans la première
moitié des années 1850, en ce qui concerne les
modalités de la formation des cellules. Il est alors établi
indépendamment par Robert Remak (1815-1865) et
Rudolf Virchow (1821-1902) que toute cellule se forme
par la division d’une cellule préexistante. Un peu
plus tard, au tournant des années 1870, l’observation
microscopique devenant plus précise, les divisions
cellulaires sont décrites chez les végétaux, puis chez
les animaux, le processus de partage des chromosomes
s’avérant remarquable de constance et d’universalité.
Au cours du XIXe siècle, la cellule s’est donc imposée
comme une structure universelle unifi ant la
compréhension du monde vivant.
Dans ce contexte, le protoplasme, la matière consti-
tuant les cellules, attire l’attention des physiologistes.
Il est ainsi au cœur des Leçons sur les phénomènes de
la vie communs aux végétaux et aux animaux données
par Claude Bernard (1813-1878) au Muséum d’His-
toire naturelle entre 1872 et 1874. Dès la deuxième
leçon, Bernard distingue trois formes de vie, mais il
ne sépare aucunement les animaux et les végétaux, au
contraire, car son but est de préciser ce qui est physio-
logiquement commun à l’ensemble du monde vivant.
Il propose donc de distinguer « trois formes de vie » :
« 1° la vie latente ; vie non manifestée.
2° La vie oscillante ; vie à manifestations
variables et dépendantes du milieu extérieur.
3° La vie constante ; vie à manifestations libres
et indépendantes du milieu extérieur. »7
La première forme de vie peut concerner aussi bien
les graines que certains invertébrés et la deuxième la
plupart des animaux, ainsi que les végétaux. La troi-
sième, en revanche, ne considère que les oiseaux et les
mammifères.
L’ensemble des leçons tend à délivrer une explication
du vivant à l’échelle du protoplasme, matière que
Bernard désigne comme le constituant fondamental
des cellules, dans lequel réside l’activité vitale.
« Le protoplasme seul vit ou végète, travaille,
fabrique des produits, se désorganise et se
régénère incessamment : il est actif en tant que
substance et non en tant que forme ou fi gure. »
« Le phénomène fondamental de la création
organique consiste dans la formation de cette
substance, dans la synthèse chimique par
laquelle cette matière se constitue au moyen
des matériaux du monde extérieur. »8
Ce sont des diff érences de constitution du protoplasme
qui permettront au physiologiste d’expliquer les diff é-
rences entre les phénomènes respectivement caracté-
ristiques des animaux et des végétaux. Bernard décrit
longuement un protoplasme vert ou chlorophyllien. Il
affi rme qu’il y a du protoplasme vert et du protoplasme
blanc, chez les animaux, comme chez les végétaux. Les
deux protoplasmes ont, selon lui, des comportements
chimiques distincts.
Les travaux de Bernard ne sont qu’un exemple parmi
les très nombreux et très signifi catifs eff orts d’une
physiologie qui tend vers une chimie biologique,
domaine qui évoluera en biochimie. Cette dernière
sera également une discipline unifi catrice du vivant,
tout en possédant les moyens de révéler et d’expliquer
des phénomènes spécifi ques.
Les développements de la génétique au début du XXe
siècle vont dans ce même sens de la construction d’une
unité des phénomènes fondamentaux du vivant. Les
travaux de Gregor Mendel (1822-1884) (publiés en
1865, ils passent inaperçus pendant 35 ans et seront
redécouverts en 1900) portaient, comme cela est bien
connu, sur le petit pois. De Vries, Correns et Tchermak
qui les révèlent travaillaient eux aussi sur des végétaux
et ce fut l’œuvre de biologistes tels que Bateson ou
7 C. Bernard, Leçons sur les phénomènes de la vie communs aux animaux et végétaux, Baillère, Paris, 1878, p. 67.
8 Ibid., pp.202-203.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
1
/
58
100%