LA LYMPHOCYTOSE POLYCLONALE CHRONIQUE B : ÉTUDE IN
publicité

MARGUERITE MASSINGA LOEMBÉ
LA LYMPHOCYTOSE POLYCLONALE CHRONIQUE B :
BIOLOGIQUES DES
LYMPHOCYïES T ET B.
ÉTUDE IN VITRO DES PROPRIÉTÉS
Memoire
Resent6 la faculte des études supérieures
de l'Université Laval
pour L'obtention
du grade de maître ès sciences (M.Sc.)
Département de Biochimie
DE SCIENCES ET GÉNIE
UNIVERSITE LAVAL
FACULTÉ
Q Marguerite Massinga Loembé, 1998
National Library
Bibliothèque nationale
du Canada
Acquisitions and
Bibliographie Services
Acquisitions et
seMces bibliographiques
395 Wellington Street
OttawaON K1AONO
OttawaON K I A W
Canada
canada
395. nie Wellington
The author has granted a nonexclusive licence aLlowing the
National Library of Canada to
reproduce, loaq distn'bute or sell
copies of this thesis in microfonn,
paper or electronic formats.
L'auteur a accordé une licence non
exclusive perniettant a la
Bibliothéque nationale du Canada de
reproduire, prêter, distribuer ou
vendre des copies de cette thèse sous
la forme de microfiche/nIm, de
reproduction sur papier ou sur format
électronique.
The author retains ownership of the
copyright in this thesis. Neither the
thesis nor substantiai extracts fiom it
may be printed or othenvise
reproduced without the audior's
permission.
L'auteur conserve la propriété du
droit d'auteur qui protège cette thèse.
Ni la thèse ni des extraits substantiels
de celle-ci ne doivent être imprimés
ou autrement reproduits sans son
autorisation.
Canada
La lymphocytose polyclonale chronique B (LPCB)est un dCsordre hématologique non malin
a des lymphocytes B du sang pCnphérique,
récemment associé au virus Epstein-Bair (EBV). Le but de ce projet de recherche éîait de
caractériser in vin0 les lymphocytes B et T chez des patientes atteintes de LPCB. L'evaluation
caractérisé par une augmentation de llgM
des fonctions générales des T n'a rCvélC aucune clifErence entre les patientes et des t h o i n s
nomiaux Grâce aux lignées immortalisées (LCL) obtenues par infection de lymphocytes B
avec EBV,il sera possible de dCtenniner si une dCfailIance de l'immUiit6 cellulaire spécifique
aux antigènes viraux pourrait être responsable du syndrome. De plus, l'activation des B via le
récepteur cellulaire CWO a d€montré que contrairement aux cellules normales, les
lymphocytes B isolCs de patientes ne proWerent pas en r C p o w B la stimulation. Une
corruption de la chaîne de signalisation de (3D40 pourrait donc modifier chez ces cellules la
réponse aux signaux physiologiques responsables de l'homéostasie du système immunitaire.
AVANT-PROPOS
Avec ce mémoire s'achève une Ctape importante de mon parcours acadbmique qui a et€
rnarquke de travaux, de collaborations et d'échanges enrichissants tant sur le plan scientifque
que sur le plan des relations interpersonnelies.
J'aimerais tout d'abord exprimer ma reco~aissanceenvers mes parentsTPaul et Franche
Loembé, grâce auxquels j'ai pu réaliser ces &tudes. En effet, ce sont eux qui m'ont toujours
poussée B m'instruire et & explorer les domaines qui m'intéressaient et qui, le temps venu,
m'ont fourni l'opportunité de partir au loin pour obtenir la meilleure formation. La distance
ne les a toutefois pas empêch6s de faire preuve d'un soutien assidu. Merci aussi B mes sœurs,
Sara et Laura, ainsi qu'a mon f&e, Junior, pour leur support et l'intérêt dont ils ont toujours
fait preuve envers mes progrès.
Merci ii Gouled, mon compagnon de tous les jours qui a partag6 mes hauts et mes bas, et
qui n'a jamais manqu6 de me prodiguer des encouragements et de me manifester un soutien
sans faille.
Par la suite, je voudrais exprimer ma gratitude envers une formidable équipe de
chercheurs : le Dr Noella DeslaUriers, ma directrice, ainsi que les Dr André Darveau et Dr
Robert Delage, mes codirecteurs. Tous trois m'ont confi6 un projet de recherche captivant et
stimulant. Ils ont su me wmmuniquer leur enthousiasme et partager leurs vastes
connaissances et expérience, et m'ont fourni un encadrement hors pair (que ce soit de vive
voix ou via coumer électronique depuis la Suisse). Grâce B eux j'ai, je l'espère, pu
développer mon sens critique et acquérir un vCritable esprit scientifique.
Je suis très obligCe envers le Dr Jean Gosselin au CHUL et son Cquipe, notamment Suie
Arsenault, dont l'expertise m'a permis d'obtenir des lignées lyrnphoblastoïdes. Cela a dom6
un coup de pouce non nbgligeable mon projet de recherche. Milles mercis aussi au
personnel du d6partement d'hématologie de l'Hôpital Saint-Sacrement, notamment Louise
Jacques, Maude Boucbard, Denise Bilodeau et les infimiières pour leur disponibilite et leur
aide technique.
Je ne peux manquer de mentionner tous les membres du laboratoire du Dr André Darveau
qui m'ont accueillie parmi eux avec enthousiasme :Josée et Josée. Carle, Geny, Rejean, et
dernièrement Marie-Ève. J'ai beaucoup apprécie nos discussions (scientifiques ou autres)
ainsi que les conseils et suggestions qui ont grandement contribu6 B l'avancement de mon
projet de recherche. Merci B Anaie et h d g e r pour leur aide technique et nombreux trucs.
Enfin, la réalisation de cette m a s e a Cîé rendue possible grâce une bourse que m'a
octroyee le FCAR-FRSQ. J'aimerais donc remercier cet organisme pour son soutien financier.
4
TABLE DES MATIERES
1
La IpphofybSe poIycIO118kchronique B
m . w m . ~ ~ m m ~ ~ ~ . m o ~ m ~ m ~ o m m m w m m m ~ m t ~ ~ m ~ m m m m m e m ~ m m o m m ~ ~ o o m m ~ o ~ 9
1.1
1.2
1.3
Historique et description du syndrome...................................................... .......--.....9
Uiologie de la LPBC :faits et hypothèses ...............
.................... 12
Vers une evolution maligne ? ................................................................................ 14
2 Le v h s
1s
2.1
Biologie du virus ...........................~.....................................................................
15
2.2
Histoire naturelle du virus dans I'organisme ........................................................ 1 9
23
Repense immunitaire à l'infection virale .....................
.
.
...................... .....ttt.....20
2.4
Implication d'EBV dans les maladies lymphoprolif6ratives................................. 21
3 &tude e t -tëriSatio~~ d~ I p p h ~ ~ m
3.1
Le developpement des lymphocytes B ..............
.-.....................A3
...... ....24
3.2 Rôle de CD40 dans le cycle cellulaire des lymphocytes B ....................
33
Système de culture in vitro d6pendant de CD40 .......................
.................... 29
4
Objectifs du projet de mhemhe.,,,
~
~
~
~
h
~
~
m
a
~
m
~
.
.
m
~
~
~
a
m
~
~
~
~
a
~
~
~
~
~
~
Bmomm~m~~~oooooom.m.mmmmoommoooomeommHmmmommooooo23
.
.
.
~
m
~
~
~
~
~
~
~
m
a
~
~
H
~
~
~
~
~
O
LISTE DES FIGURES
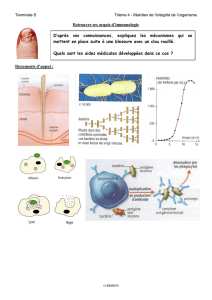
Fi-
.
1 Morphologie des lymphocytes B binuciéçs chez les patients atteints de LPCB
.
......10
Figure 2 Stmcture gCnCrale du virus EpsteimBan................................................................. 15
.
Figure 3 Organisation LinCaire du genome du virus Epstein-Barr ..........................................16
.
Figure 4 Représentation schématiqye du processus d'infection par le virus EpstebBarr....19
F
F
.
m 5 Modèle thdorique de la cascade de signalisation cellulaire dkclenchk par CD40.26
i 6. Cycle cellulaire des lymphocytes B :phase de m a t u d o n dépendante de I'antigène
...............................................................................................*...................................................28
Figaie 7. Système de culture ddpendant de CWO.................................................................. 35
F i 8. Detection de l'apoptose par test TUNEL ................................................................. 39
F i i 9. Évaluation de I'immunité cellulaire gCnCraie par stimulation mitogénique.............42
.
Figare 10 RoiXeration cellulaire r&ultant de la stimulation via CWO chez les lymphocytes
B .................................................................................................................................................
F
F
.
44
.
.
i 11 Detection de la phosphorylation sur tyrosine suite & la stimulation via CD40......46
.
i 12 Prolifdration cellulaire dépendant de la stimulation via CD40 chez une U3L issue
de ceiluies d'un témoin.... .........*......................*............................
.
F i i 13 Évaluation de l'apoptose chez une LCL suite & la stimulaîion via CW0..............50
.
Figure 14 Modèle thborique de la cascade de signalisation cellulaire dtclenchée par LMP-1
................................................................................................................................................... 57
.
Figure 15 Optimisation du test de stimulation la PHA en fonction de la concentration
ceutdaire.................................................................................................................................... 68
Figare 16. Optimisation du test de stimulation & la PHA en fonction de la concentration de
. .
mitogene...........................................**............*.............*................................
..............*..*.*..*.69
.
F ' i 17 Cinétique du test de stimulation B la PHA............................................................. 70
LISTE DES TABLEAUX
Tableau 1 :Gènes de latence du v i a Epstein-Barr essentiels B la transformation cellulaire. 18
TabIeau 2 :Tableau chique des patientes atteintes de LPCB........................................... C....C40
Tableau 3 : Évaluation de l'expression des marqueurs d'activation exprimés B la surf'e des
................................................................................................. 47
LCL. ....................
*....
Table~u4 :RCsumé des effets induits par Ia stimulation via CWO sur la prolif6ration
cellulaire chez les lymphocytes B et les LCL issues de tCmoins normaux et de patients. ........48
Tableau 5 :Calcul de la concentration cellulaire en fonction du nombre de lymphocytes
.......................................................................................... .*.............71
T/puits.. ........................ .
.
LISTE DES ABBRÉVIATIONS
BCR :Récepteur ceuulaire B.
CMA :Complexe majeur d'histocompatibilité.
CI'L :Lymphocyte T cytotoxique.
DMF :DimCthyIformamide.
DMSO :Dim6thylsuEoxide.
EBV ou HHV-4 :Virus Epstein-Barr.
FITC :Fluorescéine.
PE :P h y c o é ~ n e .
Ig :Immunoglobuline.
IL :Interleukine.
LCL :Lignk lymphoblastoïde.
LDH :Lactate déshydrogénase.
LPCB :Lymphocytose polyclonde chronique B.
MTT :Bromure de 2,5diphCnyl tétrazolium 3(4,5diméthylthiazol-2-yl).
n. :Nonnale.
NBT-BCIP :(bleu de chlorure de nitrotétmzoliurn)-(d de ptoluidine Ibmmo4chloro-3indolylphospbate)
PBMC :Cellules mononucléées du sang périphérique.
PBS :Tampon de phosphate salin.
PHA :Phytohhag glutinine.
PVDF :Membrane de bifluorure de polyvinyldiène
SDS-PAGE:Électrophorèse sur gel de polyacrylamide en présence de dodtcyl de sulfate de
sodium.
TCR :Récepteur ailulaire T.
INTRODUCTION
1
1.1
La tymphocgtov polycbde cbroiiqoe B
Historique et description du syndrome
Une lymphocytose consiste, comme l'indique son mm, en une augmentation du nombre
de lymphocytes B ou T en circulation au-dessus de la valeur Limite de 4~10%(Wïiams,
1990). On distingue deux fomes de lymphocytoses :
Une forme passagère, souvent observCe B la suite d'un accident ou autre type de
traumatisme, ou encore à la suite d'une infection M e .
Une forme durable, plus rare, et, en ce qui concerne les lymphocytes B, souvent reli6e un
syndrome lymphoprolif6ratif maiin comme la leucdmie LymphoIde chronique. Dans ce
cas, la lymphocytose rCsulte de l'expansion
d'un clone de lymphocyte B. On parle alors de
prolifération monoclonale (Hanto et al.. 1995;Williams, 1990).
Pour distinguer les deux formes, il suffit de procéder B l'examen des chaînes légères w et A
des immunoglobulines exprimCes B la surface des lymphocytes B (Hanto et al., 1995;
Perreault et al., 1989). Dans le cas d'une prolif'ration monoclonale, les cellules originaires du
clone initial prbsentent toutes le même réarrangement de leurs gènes des imr,i,iunoglobulines.
Elles expriment donc préf6rentiellement un type de chaûie légère.
En 1982, Gordon et collaborateurs deaivent pour la premi&refois un syndrome observe
chez trois patientes et consistant en une lymphocytose B polyclonale et durable (Gordon et al.,
1982). Au cours des annees suivantes, plusieurs autres chercheurs allaient rapporter des
observations semblables, contribuant B dresser un tableau clinique plus complet des personnes
affectees et du syndrome, qui prend le nom de lymphocytose polyclonale chronique B (ou
L m ) . En voici les faits saillants :
La LPCB est rencontrée presque exclusivement chez des femmes d'âge variable et
fumeuses (à ce jour, seulement cinq hommes ont été diagnostiqués (Cdlet-Bauchu et al.,
lm Troussard et al., 1997a)). La majorité des patients possèdent au moins un d è l e
--DR7
du CMH (AgrawaL et al., 1%
Troussard et al.,19%).
Tous présentent une lymphocytose, variant d e 4 à 175xl09/1, due à une augmentation de
leurs lymphocytes B (A-wal
et al., 1994). Cette augmentation est polyclonale, comme
l'indique un rapport KAnormal des chaînes Légères de leurs immunoglobulines (Gordon et
al., 1982; Troussard et al., 1994). Dans trois cas isolés cependant, l'augmentation était
oligoclonale (Chan et al., 1990; Delage et aC., 1992). Ces lymphocytes B présentent une
morphologie caractéristique : leur cytoplasme est abondant, mais surtout, dans une
proportion variant de 1% à des extrêmes de 25%, ils sont binucléés (Mitîerer et al., 1995).
Ce phénomène n'étant pratiquement jamais observe chez des personnes saines, la détection
de ces cellules binucléées dans les frottis sanguins est d'ailleurs un dément important du
diagnostic.
Fi-
1. Morphologie diif6rentielle des lymphocytes B chez les patients atteints de LPCB.
Le taux de lymphocytes T et leur répartition entre popdations CDBc et C M + sont quant
eux normaux (Delage et al.,1992, Troussard et aL, 1994).
La lymphoproIifi5ratiionest invm-ablement accompagnée d'une sécrCtion d ' I N supérieure
5 la norrnaie (cette dernitre variant de 15 B 3.0 gli). fi paraiI&le,certains ont rapporté une
diminution des taux d'IgG et d'IgA sériques (Gordon et al., 1982; Pemadt et d.,1989;
Troussard et al., 1994).
Ces obse~ationspourraient indiquer une incapacité des
lymphocytes B & e f f m le changement de classe des immunogIobdines et
sécréter
d'autres types d'anticorps que les IgM. Une étude fait aussi mention d'une augmentation
de la concentration de CD21 &que (Larcher et al., 1995). Toutdois cette obsewation ne
porte que sur une seule patiente et n'a pas tîé c o n f i é e par d'autres chercheurs.
Une association a Cgalement été Ctablie entre la LPCB et la présence de rtarrangements
entre le g h e bcl-2 et la chaîne lourde des immunoglobulines (Delage et al., 1997; Delage
et al., sous presse).
Ces réarrangements, qui ont pour conséquence de diminuer la
susceptibilité des lymphocytes B B la mort par apoptose, sont aussi obsmés chez des
individus sains. Cependant la fmuence en est grandement augmentée (environ 100 fois)
chez les personnes atteintes de LPCB.
Enfui, des ttudes plus récentes, portant sur 21 patients, tant hommes que femmes, ont
permis de mettre en Cvidence chez 19 de ces derniers un phénomène de condensation
prbmaturée des chromosomes lors des phases Gl et G2 du cycle cellulaire, ainsi que la
prkence d'un marqueur chromosomique surnuméraire : isochmmosome 3q+i (3q) (Callet-
Bauchu et
(11.
1997; Troussard et al., 1997b). Cette dernibre anomalie avait dé@ CtC
observee chez une autre une patiente en 1989 (Pemauit et al., 1489).
Du point de vue clinique, les patients présentent très peu de symptômes. Certains se
plaignent de fatigue chronique et, dans approximativement la moitié des cas, une
splbnomégalie md6rée a pu être obsewée. Cette absence de symptômes peut expliquer la
relative met6 du syndrome :les patients ne sont pas d6tectés car ils n'ont aucune raison de
consulter un médecin. De 1985P 1997,38 cas seulement ont été rapportés dans la Iittérattm
(Callet-Bauchu et al., 1997; Troussard et al., 1997a). Dans la seule région de Québec,
toutefois, une quinzaine de cas ont dCjà 6t6 diagnostiquCs dans le cadre de ce projet.
1.2 Etiologie de la LPBC :faits et hypoth&ses
P a . toutes les études mentionn&s, plusieurs ont tenté d'éclaircir les causes possibles
de la lymphocytose. En premier lieu, un lien possible entre la cigarette et le syndrome a Cté
avancé. En effet, on a constat6 chez une patiente un recul de la lymphopmW6ratio, et une
normalisation du taux d'IgM sérique lorsque cette dernihe a cessé de f m e r (Carstairs et al.,
1985). Ceaains ont ainsi spécuiC que la cigmeüc provoquerait soit l'expansion d'une
population B monoclonale minoritaire (Chan et d,1990)' soit la stimulation chronique d'une
infection virale des lymphocytes B causant 6gafement leur prolif6ration (Mïtterer et d,1995).
Cependant, certains patients sont non-fmeurs (Troussard et d.,1997b). De plus, si le tabac
peut présenter un facteur de risque, il n'explique pas l'origine de la LPCB.
D'autres font plutôt allusion & une prédisposition gCnCtique chez les patients pour
expliquer l'origine du syndrome (Troussard et al., 1994). Ii est en effet intrigant que la
majorité des personnes atteintes possèdent l'allèle HLA-DR7, si l'on considère que la
fréquence de ce dernier n'est que de 26 96 au sein de la population nord-americaine normale
(Perreault et al., 1989). De plus, comme le montrent les résultats publies récemment et
mentiom6s précédemment, ces patients présentent presque tous la même anomalie
caryotypique. M n , le fait que des lymphocytes binucléés aient été observés chez des
membres de la famille de deux patientes (Troussard et al., 1994), ainsi que le diagnostic récent
de deux sœurs jumelles toutes deux atteintes de LPCB (Canet al., L996), semblent confirmer
1' hypothèse de la cause ghétique.
Une autre piste suscite 6galement l'intérêt des chercheurs,
savoir l'implication
possible d'un agent viral. Le contrôle immunologique efficace d'une infection demande une
présentation adbquate des antigènes viraux via le CMH par la cellule infectée afm qu'elle soit
reconnue puis éliminée. Or, il a Cté dCmontr6 que 1'ailèIe HLA-DR7 etait impliqu6 dans une
présentation dCfectueuse de certains gènes viraux, permettant aux cellules infectées
d'&happer au contrôle des lymphocytes T cytotoxiques (Aimarri et al., l m ) . Ainsi, par
analogie,une infection virale chronique mal contrÔl& par le système immunitaire pourrait être
a la source de la lymphocytose persistante observe dans la LPCB. Les soupçons se sont
surtout tomCs vers le virus Epstein-Barr (EBV)en raison de son implication fréquente dans
des syndromes Iymphoprdif&atifs M n b ou malins, spçcidement chez les individus
immunodeficients (Tosato et al., 1995). Ce Wus a en effet la p@té
d'infecter les
lymphocytes B et de les rendre résistants B l'apoptose (Gregory et al.. 1991). Les premittcs
tentatives pour dCtecter la présence du virus par microscopie électronique
a par
études
~Crologiquesn'ont pas domg de résultats (Gordon et al., 1982). En 1992, puis en 1995,
l'utilisation de I'hybridation in situ et de l'ampüfkation par FCR a permis de d&tecterles
gènes viraux de latence-etlytiques chez une patiente, indiquant une infection chronique (Chow
et al., 1992; Mitterer et ol., 1995). Ii s'agissait toutefois de deux cas isolés, sans contrôle
concomitant chez des individus sains-
Dans notre laboratoire, d'antres études ont été
effoctuees en utilisant une méthode d'amplification plus sensible: le PCR
amorces
emboîtees (deux Ctapes successives d'amplification avec des amorces distinctes). Elles ont
permis de mettre en Cvidence la présence du vims chez sept patientes, alors qu'il demeurait
indétectable chez autant de témoins normaux (Ryckman et al., en soumission). Ces résultats
indiquent donc que le vims serait associb B la LPCB. L'Ctat courant des ~ o d s s a n c e sur
s les
causes du syndrome pourrait donc être résume daos ce-
phrase d9Agrawal:
uEBV may be an essential trïgger, or step, in a multifactorial process that leads to PPBL
(LPCB) in genetically predisposed individuab (Agrawal et al., 1994)
En résumé, les causes invoquées pour expliquer la lymphocytose peuvent être regroupées
en deux catégories :
Une production accrue de lymphocytes B La suite d'une stimulation chronique ou d'une
infection virale chronique.
Une durée de vie prolongée causée par une rCsistance B l'apoptose résultant d'une
infection virale, d'une surexpression de bcl-2 ou d'un défaut intrinsèque (ie :genttique)
chez les lymphocytes B.
13
Vers une Lvolution maligne ?
Le souci actuel des chercheurs est surtout de déterminer si la LPCB pourrait avoir une
6volution maligne, c'est & dire donner lieu i un pn>lif&atbnmonodonaie. Le rapport des
chaînes 16gères des immunoglobulines K et A porte B croire que la lymphocytose est
polyclonde. Cependant, comme semble indiquer l'expansion de lymphocytes B présentant
des réanangements inhabituels entre la chaîne lourde des immunoglobulines et le gène bcl-2
(Delage et al., 1997,Delage et al, sous presse), un risque demeure qu'il existe une souspopulation de lymphocytes B monoclonale (ie : constituée de cellules qui pdsenteraient le
même réarrangement de leurs gènes des immunoglobulines).
Chez trois patientes, des
analyses Southem ont d'ailleurs permis la dbtection de telles cellules (Chan et al., 1990;
Delage et al-, 1992). Toutefois chez la majorité des patients atteints de LPCB. ces souspopulations monoclonales demeuraient inddtectables. En outre, des tentatives visant & mettre
en évidence la pdsence de proliferations monoclonales avec des mCthodes de ddtection plus
sensibles comme l'hybridation in situ et le PCR n'ont pas eu de succès (Agrawal et al., 1%
Troussard et al., 1994). Le cas d'une patiente atteinte de LPCB puis ayant dCveloppé un
lymphome malin 19 ans plus tard a W rapporté (Roy et ai., lm).Cependant, ni la prCsence
des réarrangements bcl-2 caractét+stiques, ni celle du virus Epstein-Barr n'ont pu être
détectées dans les cellules du lymphome, a qui empêchait d'établir un lien avec le syndrome.
11 reste qu'un suivi régulier des patients et des études plus approfondies seront nécessaires aflm
d'karter dkfinitivement tout risque d'6volution maligne dans la ISCB.
Les similitudes existant entre la LPCB et les maladies lymphoïdes reliees B EBV. de
même que l'association mise en évidence avec le vinis, indiquent que a dernier pourraitjouer
un rôle important dans l'btiologie da syndrome. Il serait donc int6ressant de scruter plus en
détails l'implication possible d'EBV dans la LPCB et de se pencher sur le rôle jou6 par le
système immunitaire des patientes dans le contrôle d'une Cventuelle infection vide.
2.1 Biologie du virus
En 1964, Epstein, Barr et Achong obsenmt par rnicro~copieélectronique la p ~ & e
d'un virus dans une lignée ce11uIaire qu'ils ont Ctabiie 3k partir de cellules isolées d'un
lymphome de Burkitt (Epstein et d., 1964). Ayant une morphologie caractéristique de la
famille des Herpesvitidae, ce nouveau virus prendra le nom de ses dCcouvreurs :le virus
Epstein-Barr (EBV)ou HHV4. EBV est classé dans le genre l ~ p h o ~ t o v i r du
u sfait de
son spectre d'hôte restreint in vitro aux lymphocytes B de deux primates :l'homme et le singe
tamarin (Marelle et al., 1993). Il se distingue aussi par sa capacitt
infecter ces cellules de
façon latente provoquant leur proiif6ration et leur immortaIisation la fois in vitro et in vivo
(Gregory et al., 1991; Robinson et al. 1982). Toutes ces propnCtCs lui confèrent d'ailleurs un
potentiel oncogénique. Ii existe deux sous-types de virus Epstein-Barr, soit EBV-1 et EBV-2,
se distinguant principalement par leurs g&nesde latence n u c l é a h ou ebna (Khanna et ai.,
1995).
n-de
ADN double brin
tégirment
F
i 2. Stmctun générale du virus Epstein-Barr.
EBV est un virus envelopp6 de 120 B 22û nm de diamètre (Dewhwst, 1997). dont la
structure détaillée est ccprésentée h la figure 2. L'enveloppe virale est recouverte d'une
glycoprotéine majeure: la gp3501220 (Ki& et al., 1990).
L'entrée du vins dans le
lymphocyte B résulte de l'interaction entre cette glycoprotéine et la molécule de surface
-1,
normalement récepteur du fragment C3d du cornplCrnent (Marelle et al.. lm).
F
m 3. Organisation linéaire du génome du Vuus Epstein-Barr
b génome viral contenu dans la capside est constitué d'un unique segment d'ADN
bicatknaire de 172 Kb. L'organisation du gCnome d'EBV, représentee & la figure 3, est bien
connue car il a et6 complètement séquenc6 en 1984 (Baer et d,19M). Il est constitué d'un
fragment délimité par des séquences terminales rbpétées ou TR, qui permettent la
circularisation du génome sous forme d'€pisome (Marelle et d,1993). En outre, il comprend
des séquences internes repétées en tandem ou IR, la plus importante €tant IR1 ;deux origines
de réplication lytiques ou Orilyt ainsi qu'une origine de réplication latente ou OriP. La
fréquence des rCpCtitions des séquences IR varie selon les souches virales, ainsi la dimension
de certaines protéines comme EBNA-LP encodée par IR1 differe parmi les souches d'EBV
(Kieff et al., 1990).
Lorsque le virus se lie
son récepteur viral, l'enveloppe fusionne avec la membrane
cellulaire, la capside est libér6e dans le cytoplasme et le genome viral est transporté vers le
noyau où il se N c u l d s e . A ce point est etabii un des deux types d'infection virale:
l'infection lytique ou l'infection latente. Au cours de l'infection lytique, EBV exprime la
totalit6 des 84 protCines encodées par son gCnome (Farrell, 1995). menant l'assemblage et la
production de virions libérés par bourgeonnement.
La synthbe macromoléculaire de la
17
cellule hSte est inhibée menant B sa mort (Kieff et d,1990). Lors de la phase d'infection
latente, le ghome viral est maintenu dans le noyau sous fonnt Cpisomique ou, pluo rarement,
intégré dans l'ADN cellulaire (Kieff et ui., 1990). Un nombre constant de 2 & 5 copies du
gCnome dTEBVest coaservC dam la cellule ('ïhorley-Lawoon et cil, 1996). Seule une quantité
restreinte de gènes est exprimée :six antigènes nucl6aires ou EBNA-1,2,3,4,6 et EBNA-P.
trois protéines trans-membninaires ou LMP-1,2A, et 2B; deux ARN nucléaires non codants
ou EBER-1 et 2 (Famll, 1995).
L'infection latente d'un lymphocyte B provoque sa
transformation :la synthèse cellulaire d'ARN et d'ADN est activée, de même que la secrétion
d'imrnunoglobulllies, I'expression des marqueurs d'activation et la division cellulaire (Kbanna
et al., 1995). Enfin, la cellule devient résistante iî la mort cellulaire programmée ou apoptose
du fait de l'activation de gènes anti-apoptotiques comme bcl-2 (Gregory et al. 1991). Des 11
gènes de latence exprimks, six sont considCtés essentiels A la transformation cellulaire et sont
décrits au tableau 1. Comme mentionne précédemment, ce processus donne naissance in vitro
a une lignée immortalisée ou lignée lymphoblastoïde (LCL). Lorsque le virus infecte un
lymphocyte B in vivo ou in vitro, l'infection latente est préferentiellement etablie. Cependant,
il arrive parfois que le cycle réplicatif soit spontan4ment enclench6 la suite de rnCcanismes
actuellement mal connus, mais qui dCpendraient de la cellule h6te (De-Thé, 1982; ThorleyLawson et al., 1996).
18
Tablesa 1:Ghes de latence du virus Epstein-Barr essentielo h la ttansfomation cellulaire.'
I Gène
Description
1 EBNA-I
MolCcule liant l'ADN, 76 kDa, toujours exprimte.
I
1
I
1
1
Fonctions : lie OriP, permet la dpfication du gdnorne et son maintien sous
forme &pisornique.
Localisation :noyau
Traas-activateur, 52-5 B a .
Fonctions :s'associe avec la proSie cellulaire liant l'ADN RBJ-K, provoque
la surexpression de LMP-1 et LMP-2,des EBNA,de la pmté'ie d'activation
celluhire CD23 et de CD21. Nécessaire à I'initialisation et au maintien de la
transformation cellulaire.
Locatisation :noyau
RotCine de rCgulation ? 89 KDa*
Fonction :impliqué dans l'initialisation de la transformation cellulaire.
Localisation :noyau.
Protéine de régulation, 109 KDa.
Fonction :rCgule l'expression de CD21 et de LMP-1.
Localisation :noyau*
EBNA-LP
Rotbine de taille variable selon l'isolat viral.
Fonctions : impliqu6 dans la croissance des cellules transformées par le virus.
Liaison de p53?
Localisation :noyau.
Trans-activatew, 63kD.
Fonctions : active le facteur de transcription NF-KB,cause la surexpression des
marqueurs d'activation cellulaires CD40 et Fas (CD%),
des molécules
d'adhésion LAF-1, LAF-3, ICAM-1 et de la molécule anti-apoptotique %cl-2.
u
t comme onconène iorsaue traasfecté chez les ceUules C~ithéhies.
I
1
Localisation :Membrane cytoplasmique.
D'après Baer et al., 1984;Devergne et al., 1998; Farrell et ai., 1995; Khanaa er d.195 et Kieff et ai.. 1990.
22
Histoire naturelle du virus dPns l'organisme
Le virus Epstein-Barr est gCnCralement transmis par la salive -et, beaucoup plus
rarement, à la suite d'une transfusion sanguine ou d'une transplantation d'organe (De-=
1982). Pendant l'enfance, le virus est souvent contracté de fagon asymptomatique (Marelle et
aL,lm)
alors que plus tard, la primo-infection peut rtsulter en une mononucléose infectieuse
dans 30 & 50 % des cas (Hanto, 1995). Il est communément admis que les cellules épithéliales
de l'oropharynx, permissives pour le cycle lytique d'EBV, sont le site primaire de l'infection
et de la production de particules Wales (Wday et d,1988;
Ki&
et al.. 1990). La
pénétration du virus dans ces cellules se ferait via une molécule analogue à CD21 (Young et
al., 1985). Au cours de leur passage dans les tissus lymphoïdes pharyngés, les lymphocytes B
sont idectks de faqon secondaire
l'état latent.
Toutefois, grâce
l'am6Iioration des
techniques de ddtection in situ du virus au cours des dernières années, ce rCmrio est
maintenant wntestt. Ainsi, les lymphocytes B seraient l'unique site de l'infection et la source
de la production virale (Karajamis et d,1997; Niedobitek et al., 1994, Thorley-Lawson et
al., 1996). Une synthèse de ces deux conceptions de la primo-infection est présentee B la
figure suivante.
Infection prirnsfIG de 1'6pilirilimnbuccal
L)mrgbocpe B
Figure 4. Reprdsentation schbmatique du processus d'infection par le virus Epstein-Bm.
Figure adaptée de Dewhwst, 19W. CTL :lymphocyte T cytotoxique, Lat 1, II ct II : différentes formes de latence
du virus définies plus loin dans le texte-
Quelque soit le mCcanisme exact de l'infection initiaIe, les lymphocytes B infectés par
le virus à 1'Ctat latent sont activé&et proWrcnt de manière polyclode. Cependant, leur
potentiel de croissance illimita est contr61C principalement grâce B l'action des lymphocytes
T cytotoxiques (CL)
de l'h8te. permettant de réduire le nombre de aliules infectées Le
virus persiste tout de même de fapn latente chez 11106 lymphocytes B dans l'organisme chez
95 % de la population :c'est M a t de porteur sain @breUeet d.,1993; Tosaio et al., 1995).
Occasionnellement, il y a réactivation de l'infection dans les cellules constituant le réservoir
viral, que ce soit les cellules Cpithéliales ou lymphoïdes, et production de v h s dCtcctab1e dans
la d i v e (ThorIey-Lawson et al.. 1996, Tosato et al., 1995). Chez une personne normale, il
s'établit donc un ttat d96quilibreentre la production de nnis d'une part et le contrôle exercé
par l'immunité cellulaire de l'hôte d'autre part.
23
Réponse immunitairr B l'infection virale
Pendant la primo-inféction, l'hôte, devant contr8ler la fois la production de virus et
l'expansion des lymphocytes B infectés B l'ttat latent, dCveloppe tout d'abord une réponse
humorale dirigée principalement contre les prot6ines lytiques du virus.
Ainsi l'examen
skrologique met en évidence l'apparition d'anticorps (IgM puis IgG) dirig6s en premier lieu
contre les antigènes tardifs de capside VCA (vual captid antigen), puis contre les antighes
tardifs de membrane MA (membrane antigen cornplex) et précoces EA ( e d y antigen). Les
anticorps spécifiques aux protéines de latence EBNA n'apparaissent qu'après 1 B 3 mois.
Seule la repense dingee contre VCA persiste pour toute la vie B un niveau stable (Marelie et
al., 1993;Tosato et d,1995). L'immunite humorale spécifique B EBV n'est toutefois pas très
efficace (Khanna et al., 1995) et le contrôle ultime de l'infection est assuré par l'immunité
cellulaire. Celle-1%est surtout constituée de lymphocytes T cytotoxiques CD8+et d'une faible
proportion de CD@.
D u fait de l'utilisation de LCL comme cibles dans les
tests de
cytotoxicité, l'immunité cellulaire caractérisée lors de la primo-infection est surtout celle qui
est dingee contre les antigènes de latence du virus. Elle concerne surtout les protéines EBNA-
3, 4 et 6. Une faible réponse est dirigCe contre les antigènes lytiques BZLF-1 et BHRF-1
(Rickinson et d.,1997).
La réponse cellulaire m6moire destin& B freiner la profifCration des lymphocytes B
infectés de f q n latente chez un porteur srM du virus EBV est mieux dbfiaie* Les effecteurs
majeurs de cette réponse sont des lymphocytes cytotoxiques T CD8e -ques
aux
antigènes de latence exprimés par les cellules infectées (Khanna et d., 1995; R i c b o n et d..
1997). Cependant tous les a n t i g h s viraux ne sont pas reconnus avec la même efficacitéAinsi les determinants antigéniques de EBNA-3, 4, et 6 sont les cibles préf6rentieUes des
C L EBNA-2, EBNA-LP et LMP-1n t suscitent que des réponses occasiomeiles, tandis que
EBNA-1 n'est pas reconnue par l e système immunitaire (Rickinson et aL. 1997). Il s'agit d'un
phénomène d'immunodomhanceentre les €pitopes.
2.4
Implication d'EBV dans les maladies 1ymphoproIifiratives
EBV a un fort potentiel oncogenique du fait de sa capacitk
transformer les
Lymphocytes B, et seul le contrôle exerce5 par le système immunitaire de l'hôte permet
d'assurer l'etat d'tquilibre obsend chez les porteun sains du virus.
Comme le résume
Khanna (1995). l'apparition de tumeurs reliées & EBV sera donc dbpendante de
l'immunocompttence de l'hôte et des antigènes viraux exprimCs par les cellules infectees.
Chez les individus immunod6primés comme les personnes atteintes du VIH, a l l e s
ayant reçu une transplantation d'organes ou atteintes du syndrome lymphoproLif6ratî.f relit5 au
chromosome X , les lymphomes dCveloppés sont associés à EBV
près de 100 k. Ces
lymphomes r6sultent en gen6ral de l'expansion polyclonale de lymphocytes B exprimant les
11 gènes de latence du virus (ou latence III) (Khanna et al., 1995). Leur régression peut être
obtenue en diminuant le traitement immunosuppresseur chez les greffés ou en procedant a une
greffe de CïL autologues spécfiques % EBV (Tosato et d.,1995). Cela illustre donc bien
l'importance du rôle jou6 par le système immunitaire pour prevenir ces prolif'érations
opportunistes.
D'autres maladies sont également associées B EBV chez des individus apparemment
immunocompétents. C'est le cas du lymphome de Burkitt endbique (associ6 B 95 %). du
carcinome nasopharyng6 (associe h 100 %), et de la maladie de Hodgkin (associé à 40 %)
(Khanna et al., 1995). Cette fois-ci, un phénomène d'bvasion du système immunitaire serait à
-
la source de ces lymphopliférations~ En effet, il est possible pour EBV de moduler
l'expression de ses gènes de Latence, donc d'bviter la reconnaissance par les CIL morleyLawson et al., 19%; Tosato et al., 1995). Ainsi les ceiîuies de lymphome de Burkitt
n'expriment que le gène e b ~ - (ou
I latence 1) tandis que dans le cas de la maladie de Hodgkin
et du carcinome nasopharyngt, seuls les produits des gènes ebna-2, h p - f et Zmp-2 sont
détectés (latence II) (Khanna et al., 1995; Riclchson et ui., 1997). EBV ne semble cependant
pas être le seul coupable car les cellules de lymphome de Burkitt présentent invaziablewnt
des translocations de i'oncogène c-nryc causant sa surexpmssion (Tosato et al., 1995).
Toutefois le virus, qui fournit un avantage prolif6ratif aux lymphocytes B tout en les rendant
résistants B I'apoptose, pourrait favoriser I'émergcnce de clones malins et mener au
développement de prolif6rations monoclonales (Marelle et d,1993).
Il est possible que des mécanismes analogues soient responsables de la lymphocytose
polyclonaie B.
Ce syndrome met en effet en jeu une prolifération polyclonale des
lymphocytes B accompagnée par l'expression d'EBV comme en témoigne la détection de
I'expression des gènes ebna-I et Imp-1 chez les patientes (Rychan et ai.. 19%).
On ne sait
cependant pas si d'autres @ries de latence sont exprimés chez ces patientes. Cela laisse ainsi
la porte ouverte & trois mCcanismes impliquant EBV pour expliquer la LPCB :
Un défaut des lymphocytes T cytotoxiques s-ques
B EBV les empêchant de
reconnaître les cellules infectées.
Alternativement, un dCfaut intrinsèque a m Lymphocytes B empêchant une présentation
admate des antigènes viraux et donc leur reconnaissance.
Enfi, une modulation de l'expression des gènes de latence d'EBV dans les cellules
infectbes leur permettant d'échapper B la reconnaissance du système immunitaire et
d'éviter la cytolyse.
3
Étude et camct&isation des iymphocytesB
Comme indiquC au début de cette introduction, plusieurs auteurs ont tenté de
caractériser les lymphocytes B de patientes atteintes de LPCB que ce soit du point de vue
genetique ou morphologique. Toutefois, personne ne s'est encore attarde aux propri6tés
fonctionnelles de ces ceilules. Pourtant une incapacité B répondre aux signaux physiologiques
gouvernant l'hom6ostasie du système immunitaire pourrait expliquer la lymphocytose
(Thorley-Lawson et al., 1996). De plus, certains indices, comme I'apparente inaptitude des
lymphocytes B de patients ii aTectuer le changement de classe (Gordon et al.. 1982; Perreauît
et d.. 1989; Laicher et al.,
1995)' indique que ces derniers pourraient en effet differer
fonctionnellement des ceilules de personnes normales. 11 serait donc intéressant d'avoir accès
à un systeme expérimental permettant d'analyser Les pmpri6tés fonctionnelles des
lymphocytes B et de comparer patient et tCmoins normaux.
3.1
Le développement des lymphocytes B
Le cycle cellulaire des lymphocytes B est un processus long et complexe qui peut toutefois
être résumé en deux Ctapes principales :
La phase de maturation indbpendante de l'antigène qui a Lieu dans les organes lymphoïdes
primaires (foie fœtal. moelle osseuse). Elle consiste essentiellement en un réarrangement
des gènes des imrnunoglobulùies (IgM et IgD), suivi de leur expression la surface de la
cellule et mène du stade de précurseur B celui de Lymphocyte B mature et naïf ( D u c ~ o s ~ ~
1997)
La phase de maturation dCpendante de l'antigène qui se dCroule quant h elle dans les
organes lymphoïdes secondaires tels les ganglions, la rate, les amygdales et les plaques de
Peyer.
A cette Ctape s'effectue la sélection des lymphocytes B selon leur spécificit6 pour
un antigène donné. A la suite d'interactions cellulaks avec les lymphocytes T et les
cellules folliculaires dendritiques, s'engage chez le lymphocyte B un processus
d'activation, de proLif6ration et de diff6renciation. Cela conduit d'abord B la formation
d'un centre germinatif puis h l'obtention de cellules effectrices dotées d'une spécifïcitt
accrue pour l'antigène, soit :le plasmocyte ou le lymphocyte B mémoire @uchosai, 1997;
Zubler, 1997).
Cette dernière phase repose principalement sur L'interaction d'une molécuie de
costimulation expRm& la surface du lymphocyte B. CWO, avec de son ligand @sent P la
surface du lymphocyte T,CD154(aussi CWOL ou gp39).
3.2
Rôle de CWO dans le cycle cellulaire des lymphocytes B
Glycoprotéine trans-membranaire de 48 U), CD40 appartient B la famiUe des
récepteurs du TNF panni lesquels se retrouvent TNF-R, NGF-R, 0x40 et Fas (CD95)(Law
et al., 1996). CD40 est une protéine costirnulatrice exprimée B la s d a c e de toutes cellules
présentatrices d'antigène ainsi que sur certains autres types cellulaires comme les
éosinophiles, basophiles, et certains lymphocytes T (Van Kooten et al.,1991). Toutefois. elle
a la particularité d'être indispensable
l'activation et la dZ56renciation des lymphocytes B.
Son ligand, CD154,est une protéine de 33 B 35 k D faisant partie de la famille du TNF et que
l'on retrouve majoritairement B la surface des lymphocytes T CW*(Law er al., 1996).
L'engagement de CD40 par son ligand provoque la trim6risation du recepteur dans la
membrane cellulaire, ce qui déclenche une cascade de signalisation chez le lymphocyte B. La
nature exacte de ces voies de signalisation n'est pas encore totalement élucïdte. Ainsi
l'activation des sérine-thréonine kinases comme les MAPK (protéines kinases activbes par un
mitogène) est induite par CD40, mais leur association directe ou indirecte avec le récepteur
n'a pu être mise en évidence (Law et al., 1996). C'est aussi le cas pour les protéines tyrosine
kinases Lyn, Syk et Fyn (Faris et al., 1994; Kehry, 1996). Par contre, dans le cas de la
protéine tyrosine k a s e JAK3 (famille des Jmus kinase), une association avec la partie
cytoplasmique de CD40 a pu être Ctablie récemment 11 est de plus fort probable que cette
interaction conduise B la phosphorylation de la protéine STA'B, sa translocation dans le
noyau, et sa fixation i la séquence activatnce GAS, déclenchant ainsi la Cranscnption de
plusieurs gbes cellulaires (Hanissian et al., 199î). D'un autre côte, il a éîé Ctabii que
l'homotrim~risationde CD4û provoque l'association de la queue cytoplasmique de la protéine
avec plusieurs molécules de la famille des proteines associCes au récepteur du TNF (TRAF),
soit :TRAF2. TRAF3 de même que TRAF 5 et 6 découvertes plus rékmment (Hsiag
d.,
1997). 11 est 6galement possible que TRAF1 se lie indirectement & CD40 via TRAF2 (Lnv et
d,1996). Ces associations avec CD40 résultent en la formation d'un compIexe de
signalisation rnembranab (voir figure 5). Les TRAF ont toutes en commun un domaine
pouvant lier l'ADN ce qui pourrait leur con-
un rôle de &gulateurs de transcrîption en avai
de la cascade de signalisation(Kehry et al., 1996;Iaw et ai., 1996). Dans le cas de TRAFZ. il
a toutefois C t t démontré que la signalisation conduirait plutôt
de transcription cellulaire: NF-KB.
En effet,
iui
l'activation d'un autre facteur
mutant doté d'une protéine TRAF2
incomplète a pour effet d'abolir l'activation de NF-KBqui e
t induite par CD40 (Rothe et d..,
1995).
TRAF2 agirait par l'intermédiaire d'une dine-thréonine kinase nouveilement
identifiéecapable d'activer NF-KB, d'où son nom NIK, pour "NF-KB-inducing kinase"
(Malinin et al., 1997). C'est cette d e r n i b Ctape qui mène 5 l'expression de nombreux g h e s
cellulaires provoquant l'activation du lymphocyte B.
Activation de la trausuîptioa
Figare 5. Modèle thCorique de la cascade de signalisationcellulaire déclenché+ par CD40.
Comme mention& auparavant, CD40joue un rôle indispensable lors de la maturation
des lymphocytes B. représenté
la figure 6- Dans un premÏcr temps, l'interaction d'un
lymphocyte B nàif avec un lymphocyte T expiimant CD154 se fait lors de la reconnaissana
d'un antighe dans un organe lymphoïde secondah. Les signaux de costimdation induits par
l'engagement de CD40, en conjonction avec la sécrétion d'IL4 par le lymphocyte T,
entraînent chez le lymphocyte B la surexpression de molCcuIes d'activation (87ou L
=
,
Fas
ou CD95, CD23), la production de cytokines (GM-CSF, IL-2,iL6) et la prolifération. Ce
lymphocyte B est maintenant appel6 cmtroblaste et sa réplication conduit à la formation d'un
centre germinatif.
Par la
suiîe,
il subit des mutations somatiques des gènes des
immunoglobulines dans le dessein d'augmenter la spécificit6 pour l'antigène. Survient alors le
changement de classe de ces mêmes ghes permettant au lymphocyte B, devenu centmcyte, de
sécréter des types d'anticorps autres que I'IgM. A cette Ctape, où doit avoir Lieu la sélection
des centrocytes ayant acquis la plus grande affinitb pour I'antigène, CD40 intervient encore
une fois. Si le lymphocyte B est capable, via le récepteur cellulaire BCR, de recomaîtn &
nouveau I'antigène ayant enclenche la réponse immune, il poumi recevoir un signal de
costimulation via l'interaction avec un lymphocyte T. C'est ce qu'on appelle Le sauvetage car
dans l'alternative contraire le lymphocyte B entre en apoptose.
Ce sauvetage r€sulte
probablement de la surexpression de la protéine anti-apoptotique Bcl-X chez le lymphocyte B
qui est induite par CWO (Law et al., 1996). A la suite d'autres interactions cellulaires mal
d k f ~ e sle
, lymphocyte B se differencie maintenant soit en cellule sécrétrice d'anticorps ou
plasmocyte, soit en cellule mémoire, et il retourne dans la circulation (Duchosal, 1997).
1
fas de reconnaissancede L'Ag
Migraiion dans la zone manteau
Hypermutation somatique.
Changement de classe des Ig.
Reco~aissancede l'Ag
O
mémoire
Figum 6. Cycle ceildaire des Lymphocytes B :phase de maturation dépendante de l'antigène.
33
Système de culhue in vitro dépendant de CD40
li est maintenant courant d'utiker la stimulation via (=WO pour Chrdier le développement
des lymphocytes B in vitro (Banchereau et al., 1991). A cetîe fui,des anticorps monoclonaux
dirigés contre le récepteur sont utilisés (Faris et al., 1% Hsing et d,1997). Cependant, s'il
n'y a pas pontage de Leur portion Fc, le signal engendré par ces anticorps en suspension peut
être faible (Faris et al., 1994). Dans notre laboratoire, nous utilisons donc plut& un système
de culture bas6 sur l'utilisation de fibroblastes murins ( L A 3 transfectés avec le gène de
CD154 et capables d'activer des lymphocytes B en présence d'IL4 ( N h net al., 119%). II est
alors possible d'observer et de mesurer chez ces derniers les CvCnements induits par CWO qui
ont été décrits précédemment :
Activation et proliferation cellulaire.
Activation des tyrosines kinases.
Activation de NF-BI
Surexpression des molCcules d'activation cellulaire.
Changement de classe des immunoglobulines.
Sécrétion de cytokines et d'anticorps.
Différenciation cellulaire.
Il faut préciser que la réponse obtenue chez certaines souches cellulaires peut grandement
varier car les effets résultants de l'engagement de CWO in vivo sont dependants B la fois du
stade de maturation du lymphocyte
B et des interactions avec le milieu environnant (autres
cellules, cytokines), conditions qui ne peuvent pas être totalement reproduites in vifro. A titre
d'exemple, ib la suite d'une stimulation par CWO, une inhibition de la prolifhtion des
cellules d'un lymphome B a CtC observCe (Heath et al., 1993).
La synth8se de toutes les informations pdsentées dans cette introduction mène aux deux
hypothèses suivantes :
La LPCB pourrait résulter de l'incapacité du système immunitaire recomsu^trtet éliminer
les lymphocytes B infectés par le virus Epstein-Barr.
Les lymphocytes B de patientes (porteursdu vims Epstein-Barr?) pinraient differer
fonctionnellementde lymphocytes B normaux et ne pas répondre adéquatement aux
signaux physiologiques contribuant au maintien de l'homéostasie du système immunitaire.
Le premier but du projet est donc d'étudier l'association entre EBV et la LPCB et le rôle
joud par le syst5me immunitaire dans le contrôle de l'infection vide. Les objectifs
spdcifiques consisteront donc :
S'assurer de I'integrité de l'immunité cellulaire gCnéraie des patients grâce
un test de
stimulation mitogénique des lymphocytes T.
Étudier I'immunit6 cellulaire spécifique
EBV chez les patients en tentant d'obtenir des
Lignées issues de leurs lymphocytes B ou LCL,soit spontanément immortalisées par le
virus, soit infectées in virro. Eventuellement, utiliser ces LCL pour réactiver in Miro des
Lymphocytes T cytotoxiques (CïL) autologues spécifiques& EBV et tester l'activitk de ces
C L dirigdes contre les antigènes viraux présentCs par les L
m
Ainsi, en comparant les resultats obtenus chez les patients et ceux de thnoins normaux,
il sera donc possible de d6termiaer si un contrôle immunitaire de l'infection virale défectueux
pourrait mener & la lymphocytose.
Le deuxième but du projet est d'étudier les propriCtés fonctionnelles des lymphocytes
B chez les patients atteints de LPCB. Lcs objectifs spécifiques seront les suivants :
Étudier la prolif6ration cellulaire suite à la stimulation via CWO chez des lymphocytes B
de patients ainsi que chez des LCL dtrivées de l'infection de ces cellules par le v h s EBV.
Étudier la prolifdration cellulaire suite P la stimulation via CWO chez des lymphocytes B
de témoins ainsi que chez des LCL dérivé& de I'infection de a s cellules par le vinis EBV.
Comparer les réponses obtenues et dCterminer si une anomalie pourraitjouer un r81e dans
l'étiologie de la LPCB.
1
Examen clinique des patientes.
Les s u patientes atteintes de LPCB participant & cette Ctude ont Cîé recrutCes par le
centre d'hématologie de l'Hôpital Saint-Sacrement (Québec, QC).
Toutes ont signé un
formulaire de consentement approuve par le comité d'dthique de cette institution. Eiles ont Cîé
soumises a un examen clinique de même qu'8 des tests sanguins effecni6s par le personnel
technique du dkpartement d'hématologie de l'hôpital qui nous a gracieusement transmis les
résultats.
2
Préparation des ceIlriles
A partir du sang de patientes et de tCmoins normaux, des cellules mononuclێes du
sang périphérique (ou PBMC)ont Cté isolées par centrifugation sur Ficoll (Pha~naciaBiotech,
Baie d'Urf6, QC). Les cellules ont et€ conservées dans l'azote liquide après suspension dans
du RPMI 1640 (GibcoBRL, Burlington, ONT) additionné de 20 % sérum foetal bovin
décompl6ment6 (30 min, 56 C,Hyclone. Logan. UT) et 10 % DMSO. Les lymphocytes B
ont kt6 obtenus par sélection ntgative en faisant deux passages sur billes magnetiques
recouvertes d'anti-CD2 (Dynal. Lake Success, NY) permettant la dbplttion en lymphocytes T
selon les instructions fournies par la compagnie. Les cultures ont dté faites dans du milieu
RPMI additionné de 10 Z de s6nun fatal bovin déeompl6menté. 100 U/mI phicilline
(GibcoBRL), 100 pgkd streptomycine (GibcoBRL) sauf Lorsqu'indiquC autrement.
3.1
Stirnulation mitogCnique des lymphocytes T
Les conditions de culture conduisant B une stimulation optimale d a lymphocyfes T ont
été ddçtermhées aux cours d'essais préliminaires (voir annexe A, page 6 8 , permettant de
conclure qu'une dose de phytohémagglutinine (PH& Sigma, Oakville, ONT)de 1pgh.lainsi
qu'une +riode d'incubation de 48 heures devaient être utilisées. Pour la stimulation finale,
des PBMC de patientes et de temoins normaux ont été suspendues dans du milieu de culture et
ont été diluées à des concentratioas allant de 2 5 X 106 & 10 X 106 cellules/ml- mes ont
ensuite été cultivées dans une plaque de % puits à fonàs plats en présence de PHA (lpgfml)
pour une période de 48 heures 37OC,5 % CO, La proM6ration cellulaire a Cte Cvduée par
un test colorim6trique basé sur la transformation du MlT jaune en formazan bleu par les
enzymes rnitochondnales cellulaires. 100 pl de culture ont CtC prélevés, additiom6s de 20 fl
de M T ï (2,s mghl, Sigma) puis incubés 3 hr à 37"C,5 % CO,. 100 fl d'une solution de
solubilisation (20 46 SDS,50 % DMF,0.WN HCI) ont dois Ctt ajoutés pour permettre la
dissolution des cristaux de formazan, et la D.O. a 6té lue B 550 nm (réference 690 nm)aprés
une incubation d'une nuit à l'obscurit6. Puisque le taux de lymphocytes B chez les patientes
était plus élev6 que chez les témoins normaux, les résultats ont étt normalisés en exprimant la
prolifkration cellulaire en fonction du nombre de lymphocytes T par puits plutôt qu'en
fonction du nombre de cellules par puits. Le taux en lymphocytes T des PBMC a €tg
détermine par cytofluorom6trie en mesurant l'expression des marqueurs de surface spécifiques
à ces cellules
(CDS,CW, CD3) chez les patientes tandis qu'il a Cté estim6
normale moyenne de 70 % chez les témoins (voir annexe B, page 71).
une valeur
3-2 Établissement des K L
Des PBMC de patientes et de témoins, fdchement isol6es ou dcgelécs, ont et6
infectées avec le virus EBV type 1 (putif16B partir de la lign6e cellulaire B95.8,ATCCf CRL
1612, fourni par le laboratoire du Dr Jesn Gosselui, CHUL, QC). L'inftction s'est faite sur
culot cellulaire 1 heure B 37OC en ajoutant 1pl de solution Wale ( 1x10' unités I d ) h 1X 1 6
PBMC dans un tube h fond rond de 13 ml. Les cellules ont ensuite Cîé suspendues
une
concentration de 15 X 106 celluies/ml dans du d i e u de culture additiome de O 5 p g / d
ciclosporine A (Sandoz, Mon-,
QC), un imm~osuppresseurutilisé pour inhiber l'action
des eventuels lymphocytes T cytotoxiques m6moires spécifiques
EBV (Pelloquin et cd..
1986). La suspension ceildaire a ensuite C t t répartie dans une plaque de 24 puits B raison de 1
d p u i t s . puis incubée B 37OC,5 % CO,. Le milieu B C t t renouveld une fois par semaine les
deux premières semaines puis tous les 2-3 jours. A l'apparition des signes de orm mat ion
cellulaire telles l'augmentation de la densite cellulaire, 1'agrCgation des cellules et
I'acidification rapide du milieu (Robinson et al., 1982). le contenu des puits a ét6 trand616
successivement dans des flacons de 25 cm2et de 75 cm2.
Une fois les lignées Ctablies, elles ont CtC testées pour la detection des mycoplasmes B
l'aide de la trousse Mycotect (GibcoBRL). Les résultats s'ttant avCrés positifs pour toutes les
lignées, ces dernières ont étC soumises & trois cycles de décontamination avec l'antibiotique
BM-Cyclin (Boheringer-Mannheim, Laval, QC). L'élimination de la contamination a et6
un nouveau test via Mycotect. Par la suite, les lignées ont Cté maintenues en
culture dans du milieu Iscove (GibcoBRL) additionné de 5 % de sérum fœtal bovin
confirm6e par
décompl6ment&
4.1
Stimulation via CD40
La méthode utilisée est décrite dans Ia litîérature ( N h n et al., 1996)- Brièvement,
après purification par retrait des lymphocytes T sur billes magnétiques, les lymphocytes B ont
été cultivés à une concentration de 25x1@ cellules/puits dans une plaque de % puits A fonds
plats contenant lxlv LA5 (fibroblastes muiins exprimant b ligand de CD40 ou CD141)
irradiees par puits. Les cultures ont été effectudes dans un volume final de 20 pI de milieu
Iscove contenant 5 % de sérum foetal bovin décompl6ment6, 100 U/ml pénicilline, 100 pg/ml
streptomycine, 100 U/mi dlL-4 (Genzyme, Cambridge, MA). 5 &ml d'insuline bovine
(Sigma) et de transiferrine humaine (Sigma). La moiti6 du milieu a été renouvelée tous les
troisjours et les L A S remplades a tous les sixjours.
CeauleLAS
Figure 7. Système de culture dbpendant de CWO.
4.2
Estimation de la prolif6ration cellulaire par MIT
Des lymphocytes B de patientes et de témoin normaux ont éié stimulés pendant des
périodes variant de 1 B 9 jours dans le système de culture d d p d a n t de CWO et la
proW'ration des cultaies effecniées en triplicaîa a Cté Cvaluéc par test calorimétrique MIT tel
que ddcnt précédemment pour le test de stimulation mitogénique B la PHA.
43
Estimation de la pmIif6ration par compte cellalaire
Des cultures ont de nouveau €té f i t e s en tnplicata dans le syst&mede proliféraion
dépendant de C'Dm, mais sur des périodes supplCmentaires allant jusqu'à 18 jours. La
prolifération cellulaire a CtC estimée par compte des cellules à I'hématimètre. Lorsque les
cultures atteignaient une densité de 2x16 lymphocytes Blpuits, la concentration &ait ramenée
& la valeur initiale de 2 5 x l e cellules/puits en diluant 1 volume de suspension cellulaire (25
pl) dans 7 volumes de milieu de culture (175 pl) dans un plaque contenant des cellules LA5
fraîchement irradiées. Le facteur de dilution &ait ensuite pris en compte pour le calcul de la
quantité totale de cellules.
4.4
Detection de la phosphorylation sur tyrosine
Des lymphocytes B d'un témoin et d'une patiente ont tté stimulés 45 minutes grâce a
un anti-CD40 soluble (G28.5, 5 pg/rnl, laboratoire du Dr Walid Mourad, CHUL,QO ainsi
que 2 heures et 72 heures dans le système de culture dépendant de CD40-Un contrôle negatif
constitue de lymphocytes B non stimul6s a aussi été effectue. Les protéines cellulaires totales
ont alors Cté extraites tel que décrit dans la littérature (Lapointe et al., 19%). en utilisant un
tampon de lyse composé de U)m M Tris-HCI pH 8.0, 137 m M NaCl. 10% (ph) glyc&ol, 1%
(ph) Triton X-100,2m M EDTA,1 mM NAJO,,
1m M
PMSF,2 pM pepstatine A. Après
dosage par Bradford, 40 pg de protéines ont été séparées par migration SDS-PAGEsur un gel
contenant 10% de polyacrylamide, puis transfierrées sur membrane de PVDF. Les protéines
phosphorylees sur tyrosine ont Cté d6tectbes B l'aide d'anticorps monoclonaux m
d
IgG2k (îpghi,Upstate Biotechnology Incorpotatbd, Lake Piacid, NY), d'un anticorps
secondaire biotinyl6 (anti-IgG, A, M, Sigma, dilut 111000). puis en utilisant le s y s t è w
streptavidine-phosphatasealcaline (ICPL, Gaithersburg, MD)- NBTfBCrP (BRL, Gaithersburg,
MD) pour la révélation.
5.1 Analyse de l'expression des molécules d'activation
L'expression des rnolCcules d'activation cellulaire CWO, CD80 (ou B7-1) et CD86 (ou
B7-2) a et6 mesurée par cytofluorométrie chez toutes les LCL obtenues & partir de l'infection
de lymphocytes B de patientes et de témoins. Les cellules ont étC iavCes 2 fois au PBS et
suspendues B une concentration fmale de 3 5x106 celluledd dans du milieu de culture. 100
ml de chaque suspension cellulaire ont été additionnés de IOpg/rnl d'anticorps dirigC contre la
moMcule Chidiée, soit un anti-CDSO coup16 la biotine (Phanningen, Mississauga, ON), un
anti-0% coup16 B la biotine (Pharrningen) ou lYantitiCD40
G28.5. Après une incubation de
30 min B 4' C et un lavage au PBS, la suspension cellulaire a ttC additionn6e d'un anticorps
secondaire coup16 à la fluorescence : soit un PE-stnptavidine (Cedarlane, Hornby, ON) pour
détecter les anticorps primaires biotynylbs, ou un anti IgG, A, M-FITC (BioCan, Mississauga,
ONT) pour la détection du GZS.5. Après une nouvelle incubation de 30 min B 4' C et un
lavage, la fluorescence a CtC mesurCe par cytofluorométrie.
5.2
Estimation de la pmlif&ratioaaiidaire par EnIT
Des LCL, obtenues partir de I'infkction de cellules de patientes et de témoins, ont été
stimulées pendant des périodes variant de 1B gjours dans le système de culture dependant de
CD40 comme décrit précédemment dans le cas des lymphocytes B, toutefois sans
remplacement des cellules U.5au jour 6. La prolif6ration des cultures effectuées en Piplicata
a été CvaluCe par test colorimCtrique
MIT tel qu'indiqué pour le
test de stimulation
mitogénique A la PKA.
5.3
Evaluation de l'apoptose par TUNEL suite & la stimulation via CD40
Une LCL de témoin a €te stimulée pendant une période de 9 jours dans le système de
culture dbpendant de CWO. Aux jours 2 . 4 5,7,9, les cellules ont et6 pr6Iev€es, lavées une
fois au PBS et suspendues dans du milieu RPMI h une concentration fmaie de 1x10'
cellules/ml.
10 pl de suspension cellulaire ont ensuite t t C utilisCs pour faire un fmttis
d'environ 1 cm de diamètre sur lame. Aprts un séchage de 30 min B température ambiante,
chaque frottis a et6 fixé 2û min dans une solution de P B S 4 % paraformaldéhyde, déshydrate 5
min successivement dans de l'éthanol 50, 70, 95 et 100 46, puis conservé
-20' C
La
presence de cellules apoptotiques a ensuite pu être CvaluCe par imrriunofluorescence en
utilisant le systeme TUNEL (Boherhger Mannheim) selon le principe representee à la figure 7
et selon le protocole fourni par la compagnie.
Des cellules murina SP2. stimulées 3 heures
avec de la cycloheximide (25 pg/ml), un inducteur de l'apoptose chez les d u l e s , ont Cté
utilisdes comme contrôle positif.
ADN
F@am ?L Détection de I'apoptose par test TUNEL
Les patientes incluses dans cette Ctude. toutes fumeuses,présentaient les signes cliniques
distinctifs associCs la LPCB, comme en témoigne les données p r é s d e s au tableau 2.
Ainsi, malgré l'absence de symptômes, toutes étaient a f f i i t s d'une augmentation
polyclonale et persistante de leurs lymphocytes B. d'une augmentation polyclonale de la
concentration d7IgMsérique et quatre patientes sur six posséàaient I'allèie HIA-DR7. La
présence de lymphocytes B binucl&s a tgalement CtC constatée dans tous les cas (donnees
non présentées).
Tableau 2 :Tableau clinique des patientes atteintes de LPCB. Les tests sanguins ont été
effectxés par le personnel technique du departement d'hématologie de l'Hôpital SaintSacrement qui nous a ensuite transmis les rtsultats.
l
Patiente Age Lymphocytose X 1û% Tarn de B
(n. :0e05-0.15)
(lL8:1.5-380)
I
(-)
l
/
L
I
IgM sérique HLA-DR suivi
I
I
2
Étude de l'immunité œllrihire
La PHA est un mitogtne qui fournit aux lymphocytes T un signal d'activation et de
proLif6ration.
Toutefois, ce dernier, contrairement au signal spécifique
engendré par le
récepteur cellulak TCR,s'dresse B la majorité des lymphocytes T. La stimulation par PHA
constitue donc un indicateur de l'efficacit6 de l'immUnit6 ceiiulak g€ndrale. A la figure 9,on
peut comparer la réponse obtenue la suite d'une telle stimulation chez des lymphocytes T de
patientes atteintes de LPCB et des lymphocytes T de témoins normaux. Pour refiéter la
fluctuation existant au sein de la population nonnale, une "zone de vmiabilit6 normale" (en
pointill6s) a Cté etablie B partir de la moyenne et de 1'écart-type des réponses obtenues avec les
cellules de 5 thnoins normaux. La proLrération observCe avec les cellules de patientes se
situait dans Les Limites de cette zone normale. Cela est une attestation de l'integrité de
I'immunit6 cellulaire génerale chez les patientes atteintes de LPCB qui corrobore l'absence
d'infections associées au syndrome. Mi d'approfondir l'Cade de I'immunitC cellulaire, il a
ensuite eté d6cidC de procéder B des tests fonctionneIs mesurant la réponse cytotoxique (mL)
spkifique aux antigihes du virus Epstein-Barr.
Dans un premier temps, iI a donc Cté
nécessaire d'obtenir des LCL, ou lignbs infectées par le virus, pouvant servir de cellules
cibles ii ces tests. Les tentatives pour obtenir des lignées spontanCment infectées à partir de
cellules de patientes s'étant soldées par un &hec (données non prtsentées), l'infection s'est
faite in vitro sur PBMC avec le v h s EBV type 1qui est Le plus fréquemment rencontre dans la
population occidentale (Robinson et al., 1982). Les cellules de cinq patientes ont kt6
infectées :deux cas (PD, PM) ont permis d'obtenir des LCL &ablies, dors que dans les trois
autres cas (PS,PC,PR) les lignées ont regresst après 1 B 4 mois de culture. De même, Les
PBMC de 3 temoins normaux (TA, T19, T41) ont été infectées, toutes avec succès, a h
dT6tabLirune réponse cytotoxique contrôle. Il est à noter que le fait d'utiliser des PBMC
congelées ou fraîches, de même que d'ajouter de la ciclosporine A au milieu de culture n'a pas
semble influencer le succès de l'infection virale. Éventuellement, ces U3L pourront donc être
utilis6es pour dCteminer la réponse cytotoxique anti-EBV de lymphocytes T autologues (voir
annexe C page 72). Eh comparant la e n s e contrôle obtenue avec les CIZ de témoins
normaux et celle obtenue avec des CïL, de patientes, il sera possible de determiner si un
défaut au niveau de l'élimination des lymphocytes B porteurs du virus peut 2tre responsable de
ia iymphopmlifération.
O
2
4
6
8
10
Lymphocytes Tlpuits (XI@)
9. Évaluation de I'immunité cellulaire genérafe par stimulation mitogenique. Les
PBMC de tdmoios et de patientes, isol6es du sang par centrSugation sur Ficoil. ont été
suspendues des concentrations cellulaires variables et stimulées avec l&d PHA. La
prolifbration a étC 6valuée après 48 heures par MIT (lecture B 550 nm,réf6rence B 690 nm).
In vivo,l'engagement de CWO par son ligand CD154 dklenche chez le lymphocyte B
nomal une cascade complexe de signabation qui dsulte ultimement en l'activation et la
prolifkration de la cellule. L'6valuation de la prolifCration ailulaire dans le système de
culture dépendant de CWO est donc un moyen aiJ de vérifier in vino si un lymphocyte B est
capable de répondre B la stimulation induite par cette molécule. Chez une personne normale,
la formation d'amas cellulaires due au phénomène d'adhésion homotypique est rapidement
observable, du fait de L'activation de I'expression des m016cdes d'adhésion cellulaire (ICAM-
1, -
1 Puis, après un d&i d'envizon 5 jours, la prolifhtion des lymphocytes B est
détectable par test calorimétrique MTT et l'on obtient une courbe de croissance cellulaire
exponentielle ( N t m et al., 19%'). C'est en effet ce qui a 6té obdervt avec les cellules de
témoins normaux (figures 10A et 10B). Avec les cellules de patientes toutefois, la
prolifkration est demeurCe à un faible niveau même après 9jours de culture, comme on peut le
voir à la figure 10A. M m de dktenniner si cette diffkrence n'&ait pas le r&ultat d'une phase
de "latence" initiale plus longue chez les lymphocytes B patientes, la proliferation a tte
estimee sur une période de 21 jours par compte cellulaire direct. Ces résultats, prkentés B la
figure 2B. semblaient indiquer une incapacité des Lymphocytes B de patientes h répondre à la
stimulation via CD40,la concentration cellulaire demeurant à un niveau de base très faible
(8x10~
celluleslpuits) même après plusieurs joua de culture. Cependant, il faut noter que
prolifkration cellulaire n'est que l'aboutissement de la chaîne de signalisation induite par la
l'interaction de CD40 avec son ligand CD154 Il a donc Ct6 decide de veiifier en amont de la
cascade de signalisation si une anomalie pouvait expliquer le phdnomène observC avec les
lymphocytes B des patientes.
O
2.5
5
75
10
Jours de cd-
5
Patiente M
4
V
Ternoin46
3-
2-
1
O
5
10
20
Jours de cuIture
Fi10. Prolif6ration cellulaire résultant de la stimulation via CD40 chez les lymphocytes
B. Des lymphocytes B de patientes et de témoins ( 2 5 X 104 ceUules/puits) ont 6îé mis en
culture avec des cellules L4.5 exprimant CD154 en présence d'ILA. A) Évaluation de la
proW6ration sur 9jours par test colo~6triqueMIT. Chaque CO&
est représentative de 3
essais effectués avec les lymphocytes B issus de 3 personnes differentes. B) Évaluation de la
prolifération sur 21jours par compte cellulaire l'h6matim&re.
3.2 Évaluation de l'activité tyrosinekinase
La phosphorylation des acides amines tyrosines étant iine des activités p&occs dans la
cascade induite par CD40 (Khey. B%),un M a u t i a niveau peut indiquer une absence
totale de
signalisation via a récepteur. Afin de v&rifiercette hypothbc, des extraits de
proteines cellulaires d'une patiente et d'un témoin n o r d ont Cté d d i s & après diff6rentes
le système de cuitpre dependant de CWO. et la tyrosine
phosphorylee a Cté dbtectée ppr analyse Wattm. L'apparition de nouvelles bandes @s
45
@riodes de stimulation dans
min, 2 h et 72 h de culture (fi-
11) témoigne de la phosphorylation de plusieurs protéines
sur tyrosine. La stimulation des cellules de patientes via CWO a donc bien induit une activité
tyrosine kinase. De plus, aucune diff'rence majeure n'a W comtat& entre les patrons de
phosphorylation de la patiente et du témoin :tant I'intensit6 que le nombre de bandes obtenues
après chaque pCriode de stimulation sont comparabies. Il semble donc que la signalisation via
CD40 n'&ait pas affectte en debut de cascade chez les patientes, ce qui pennet d'affmer que
la molécule est bien capable de fuer son ligand et de transmettre le signal résultant de cette
interaction.
Figure 10. Détection de la phosphorylation sur tyrosine suite à la stimulation via CD40.
Après des périodes de s tunulation de 45 min, 2 h, 72 h, les protéines cellulaires d'une
patiente (P) et d 'un témoin normal (T) ont été extraites et 40 pg s éparés sur gel de
polyacrylamide par SDS-PAGE. Après transfert sur membrane de PVDF, les protéines
phosphorylées sur tyrosine ont pu être détectées avec I'anticorps monoclonal rgG2b~.La
mi,ption des poids molécutaires standards est représentée à gauche.
L'section latente par le virus Epstein-Barr est un processus long et complexe qui. en
provoquant l'activation, la dinCrenciation et la transformation cellulaire
CO-
au
lymphocyte B de nouvelles propriCtés fonctionneiles (Robinson et d., 1982). C'est la raison
pour laquelle la caractérisation des LCL obtenues il p&
de l'infection de cellules de
patientes et de témoins normaux a CtC entreprise en parallèle aux Ctudes effectu6eo chez les
cellules non transformbs.
Du point de vue morphologique, toutes les lign&s présentaient les
signes typiques de transformation ceUulaûes (Robinson et al.,lm):dotées d'un cytoplasme
abondant, les cellules prbsentaient des tailles et des formes variées, elles formaient de gros
amas cellulaires et, haute densité, avaient tendance B adhdrer aux parois du flacon de culture.
La cinétique de culture variait cependant au cours du temps et d'une lignée l'autre (résultats
non présentés). L'analyse de L'expression des marqueurs d'activation cellulaire B la surface
des LCL (présentee au tableau 2) n'a révélC aucune diffhnce notable entre lignées dérivées
de patientes ou de témoins, et a confirmé l'expression de CWO à la surface des cellules
transformées.
Tableau 3 : Evduation de l'expression des maqueurs d'activation exprimes ih la surface des
LCL. Les molécules d'activation ceiiulaire à la surface des LCL issues de temoins et de
patientes ont et6 dCtect6es & l'aide de I'anticorps G285 (anti-CD4û), d'un anti-CDSO et d'un
anti-CD% tous deux couplé A la biotine. La ddtection à l'aide d'anticorps secondaires couplés
Zi la fluorescence a permis l'évaluation par cytoflorom6trie du taux d'expression de chaque
molécule,
Lignée
LCL TA
CD00
CDS0 (B7.1)
CD86 (B7.2)
90%
89%
81%
LCL T41
82 %
63%
65 %
LCL TI 9
LCL PD
97 %
68 %
85 %
91 %
68 %
81 %
4.1
Rolifdration cellulaire en réponse & la stimulation via CWO
La stimulation de la lignée TA (issue d'un témoin) dans le système de culture
dkpendant de CD40 n'a pas résulté en une activation de la prolifbtion cellulaire comparable
ce qui etait obsend avec des lymphocytes B de témoin non transfo~n€s-En effet, après une
phase initiale de proW6ration. une inhibition notable de la pmlif6raticm &ait visible après 5
jours de culture (voir figure 12). Des études effectuées avec le test TUNEL, ont permis
d'associer une augmentation substantielle de l'apoptose B la baisse de proliferation observ6e
partir du cinquième jour de culture chez Ies cellules (voir figure U). Ces résultats sont
toutefois préliminaires car ils ont CtC obtenus B partir d'une L U issue d'un seul témoin et une
fois sur trois l'inhibition n'a pas pu être reproduite de façon aussi convaincante. Le même
genre de culture, effectuées avec les lign&s dérivCes de patientes atteintes de LBCB, ont
semble indiquer que ces demiès seraient insensibles
la stimulation via CWO. Ainsi
l'engagement du récepteur CWO chez ces U=L n'a pas modif16 la cinetique de croissance
nomale observée chez ces lip6es 10rsqu'eIIes Ctaient cultivées en absence de stimulation.
Ainsi, la culture dans le système dependant de CWO a résult6 en une courbe de prolif6ration
parfaitement superposable & celle qui &ait obtenue dans des conditions de culture normales
(ie :hors du système). CeIa indique donc que I'interaction de CD40 et de son ligand n'a causé
ni augmentation
ni inhibition notable de la prolif6ration normale chez les LCL issues de
patientes.
Tableau 4 : Résurnt des effets induits par la stimulation via CWO sur la proliferation
cellulaire chez les lymphocytes B et les LCL issues de témoins normaux et de patients.
Lymphocytes B mrmmcx
Lymphocytes B tPCB
LCL nor??lUza
LCL LPCB
-
+
+
+
+
*
F i i r e 12. frolif€rationcellulaire dependant de la stimulation via CWO chez une LCL issue
de cellules d'un témoin. La lignée TA (23X 10'celluleipuits) a €té mise en culture avec des
cellules L A S (lx lO'cellules/puits) exprimant CDlSQen présence d ' m . La proiifëration sur
9jours a W 6valuCe par test colorixnétrique MIT.
F ' i m 13. Évaluation de f'apoptose chez une LCL suite la stimulation via CD40. Après 2,
5 , 7 et 9 jours de stimulation dans le système de culture dependant de 0,
les cellules de la
lignée TA ont kt6 f i e s sur lames et la présence de cellules apoptotiques a étC Cvaluée grâce
au test T m .
DISCUSSION
Aux cours des deraières années, plusieurs Ctudes visant mieux caractériser la LPCB
ont été publiées et certaines, dont une effectuée dans notre laboratoire, ont 6tabli un lien entre
le virus Epstein-Barr et la maladie. Ainsi, chez les patientes la présence des gènes de latence
du virus est invariablement détectée par PCR contrahanent & ce qui est observt chez des
témoins nonnaux (Ryckman et d,
en soumission). En foi de ces résultats, nous avons tent6, B
partir de PBMC de patientes, d'établir des lignées lymphoblastoïdes spontankment infectées
par le virus, pour ainsi confmer la présence du virus, v M e r son potentiel transformant in
vin0 et obtenir des cellules cibles pour d'éventuels tests de cytotoxicité (voir annexe C. 72).
Malheureusement, toutes les tentatives pour établir ces LCL se sont avbrées infmctueuses. Il
est difficile toutefois de préciser la signification exacte de ces résultats. En effet, l'obtention
de LCL spontanément infectées par le virus est un processus exigeant dépendant nombreux
facteurs. A titre d'exemple, il faut compter sur 1'Ctat physiologique des cellules lors de leur
prélèvement (certains lymphocytes B infectes par EBV peuvent déclencher spontanbrnent la
phase lytique lorsque places en culture) (Robinson et aL, l m ) , le nombre de lymphocytes B
porteurs du virus dans 1'Cchantillon de sang utilisé pour isoler les PBMC (ce dernier devant
avoir kt6 pr6levC dans les 24 heures précédentes) (Rocchi et al., 1977)' la densite cellulaire
lors de la culture et, enfia, la fréquence de changement du milieu de culture. En outre, certains
prkconisent la co-culture des PBMC avec des cellules adhdrentes nourrici&res(Pelloquh et al.,
1986) ou encore l'utilisation d'une dose sous-mitogénique de PHA (Robinson et d.,1982)
pour reproduire in vitro les signaux de CO-stimulationpouvant influencer la transformation in
vivo. 11 serait intértssant de vérifier si ces modifications au protocole augmenteraient
effectivement nos chances d'obtenir des lignées spontanées. Enfin, s'il y a transformation, les
lignées ne sont Ctablies qu'après un minimum de 30 jours mais cela peut prendre jusqu'h trois
mois (Robinson et al., 1982). Par conséquent, la difficult6 Cprouvée
obtenir des LCL
spontanées n'était peut-être que le résultat de conditions de culture inadéquates. Cependant, il
ne faut pas négliger le fait que ces r6sultats peuvent aussi résulter d'une incapacité du virus à
transformer les lymphocytes B in vino. On peut donc supposer un defaut au niveau dcs
protCines de latence essentielles B la tranofonnation cellulaire (voir tableau 1,page 18). Ainsi,
il est prouvt que la protéine de latence LMP-1joue un rôle essentiel dans l'initiaiisation de la
transformation cellulaire (Eiiopoulos et al., 1998) et le maintien de prolif'tion celialaire
(Kilger et al., 1998) et que des mutations de cette mol6cule peuvent affecter le potentiel
transformant du virus in vitro (Hiopoulos et al., 1998). O r B partir de 1'ARN virai isolQlors
de notre! étude chez plusieurs patientes, des mutations ont été caractérides dans cette même
protéine (Ryckman et al., en soumission). Il est donc possible que les mM1cations de la
rnolCcule LMP-I aient empêch6 la ttansformation cellulaire. L'alternative pour obtenir des
LCL a et6 par consdquent de procéder in vitro
ik l'infection des
PBMC de patientes et de
témoins. Cette méthode avait pour avantage de contrôler la "qualit6" du v i m utilis6 pour
l'infection. En outre, comme l'ont prouv6 nos résultats, Les contraintes de culture étaient
diminuées grâce la possib0ilitCd'employer des cellules préaiablement congelées. NCanmoins.
un biais pourra résulter de l'utilisation ultérieure de ces Lm En effet, il est possible que la
souche ou le type de virus présent chez les patientes m
r
e de celui qui a CtC utilise pour
L'infection in vitro. Il faudra donc tenir compte de ce dCtail lors de l'interprétation des
expériences effectuées. Cette dernière rCserve mise B part, il sera possible d'utiliser les LCL
comme cibles lors de tests de cytotoxicitt visant B determiner si le systéme immunitaire des
patientes est capable de reconnaître et d'éliminer les cellules infectées par le virus.
A ce jour, l'activité cytotoxique dirigée contre EBV a suriout Cté caractérisée chez les
lymphocytes T C D S .
En outre, cette
repense
est dingée prtfhntiellement contre les
protéines de latence EBNA-3,4, et 6, du fait d'un phénomène d'immunodominance entre les
épitopes viraux. Une petite proportion de la réponse immunitaire est Cgalement assurte par
des lymphocytes T CM+, mais elle est moins bien dtfinie (Ricbson et al., 1997). Dans un
premier temps, il conviendra donc de determiner s'il y a une réponse cellulaire spécifique
EBV efficace chez les patientes. Dans l'affmative, il sera alors possible de mieux la
caractériser et de determiner sa composition en effecteurs
et CD4+. Le rôle joué par les
lymphocytes T CD4' est particulièrement intéressant en raison de l'association Ctablie entre la
LPCB et I'ailèle HLA-DR7. En effet, cet ailèle est un membre du CMH de classe II qui est
impliqué dans la pr6sentation des antigènes aux lymphocytes T CD#. De plus, il est associC B
une présentation dbficiente d'antigènes viraux dans le cadre de l'hépatite B. Or, une autre
particularité de la W n s e immunitaire spécifique B EBV, en plus du phhornbe
d'immunodominance entre Ies antighes viraux sus rnentiom6, est la restriction des épitopes a
un allèle donné du CMH. Ainsi, un Cpitope sera prCsent6 spécifÏquement par un alltle précis
du CMH (Rickinson et al.. 1997). La combinaison de ces deux facteurs peut donc impliquer
qu'une réponse assumée par M tymphocyte T CD4t contre un épitope dominant et présenté par
l'allèle FILA-DR7 serait essentielle au contrôle de I'infection virale. Une telle réponse, si eile
etait dbficiente chez les patients atteints de LPCB pourrait possiblement avoir un d e à jouer
dans 1'6tiologie du syndrome.
Le second volet de ce projet de recherche consistait B évaluer les propriétés
fonctio~ellesdes lymphocytes B chez les patientes atteintes de LPCB grâce la stimulation
via le récepteur cellulaire CD4û. Le déclenchement de la proliféation cellulaire est un effet
bien documenté résultant de l'interaction de CD40 avec son ligand CD154, et c'est le
paramèoe qui a étC utilisé dans ce projet pour Cvaluer la rkponse des lymphocytes B
stimulation.
a la
Les résultats obtenus ont clairement démontrC que, contrairement aux
lymphocytes B de temoins, la population cellulaire chez les patientes n'augmentait pas de
façon considérable, même a p r b 20 jours de culture dans le système dépendant de CD&. Cela
laissait donc supposer une absence de prolif6ration donc signalisation via CD40 chez les
cellules de patientes.
L'analyse de la phosphorylation de la tyrosine, qui est un des
kvenements pdcoces induits suite B la stimulation, a cependant dvélt que la molécule CWO
exprimée par les lymphocytes B de patientes etait bel et bien capable d'interagir avec son
ligand et de gbnerer un signal. Toutefois, cela n'exclut pas la possibilite d'une corruption de
la cascade de signalisation induite par CWO chez les patientes. C'est ce que semble indiquer
l'absence de proliftration des lymphocytes B de patientes suite à la stimulation in vitro, de
même que leur apparente incapacit6
effectuer le changement de classe in vivo, comme en
tkmoigne le taux d'IgM supérieur & la normale associC au syndrome. Ces deux CvCnements
interviennent plus tardivement dans l'activation ceilulaire et, selon les connaissances actueues,
seraient préc6dés de l'augmentation de la transcription de certains gènes cellulaires. Or ces
6vvénements induits par CD40 résultent de voies de signalisation complexes et variées (voir
figure 5, page 26) encore mal caracténsCes. Plusieurs zones d'ombre subsistent encore en ce
qui concerne les mécanismes moléculaires exacts gouvernant l'association de CD40 avec les
mol6cules de signalisation cellulains comme les TRAF. C e s t aussi le cas en ce qui a trait B
l'enchaînement des voies de signalisation résultant de ces interactions. Petit h pctit toutefois,
des études parviennent B démêIer les liens reliant les ph6nomènes induits lors de l'activation
cellulaire aux diff6mtes voies de la cascade de signalisation déclenchée par CÛ40.
L'interaction avec 'ïRAF2 serait ainsi responsabk des CvCnements précoces comme
l'expression des marqueurs d'activation cellulaire et la prolif&ration,alors que l'association
avec TRAF3 serait plutôt impliqu& dans le changement de classe des immunoglobulines
(Kuhné et al., 1997). En conclusion, il faudra donc d'abord vtntier chez les patientes quelles
sont exactement les Ctapes d'activation f9isant défaut afin de parvenir B localiser une anomalie
dans la cascade de signalisation engendrée par CWO. Ainsi, on pourra facilement vérifier la
capacité des lymphocytes B de patiente activer le facteur de transcription cellulaire NF-uB.
provoquer la surexpression de rnolCcules d'activation et
effectuer le changement de classe
des immunoglobulines. Cependant, à ce point, rien ne permet de distinguer quel Clément, du
récepteur ou des mol6cules de signalisation, pourrait être dkficient.
Chez les patientes atteintes de LPCB, la présence d'une population de lymphocytes B
porteurs d'EBV et l'expression accrue des g&nesde latence viraux (tel qu'indiqué par PCR)
peut également influencer la
repense
des cellules & la stimulation via CD4û. Beaucoup
d'intkrêt a CtC accord6 rCcemment aux interactions prenant place entre ce récepteur et les
protbines du virus EBV,plus particuli+rement la p t 6 i n e de latence LMP-1. En effet, LMP-1
partage beaucoup de similitudes avec les molCcules de la famille des récepteurs du TNF dont
fait partie C M . Ainsi, LMP-1 est elle aussi une protbine membranaire qui, en s'agrkgeant,
provoque le recrutement des TRAF? menant à la fornation d'un complexe de signalisation
cellulaire (Gires et d,1997; Kilger et al., 1998). Ce complexe est vraisemblablement à
l'origine de l'activation du facteur de transcription cellulaire NF-&, ainsi que de la
surexpression de marqueurs d'activation, parmi lesquels CD40 et Fas, de mol6cules
d'adhésion et d'autres protéines cellulaires (voir tableau 1, page 18) (Devergne et al., 1998).
LMP-1 joue donc un rôle essentiel dans l'initialisation de la transformation et la prolifération
chez les lignees lymphoblastoïdes (Eliopodos et al., 1998; Kilger et al.. 1998). Une
distinction fondamentale existe cependant entre LMP-1 et les molécules de la famille des
récepteurs du TNF au niveau structural : la protéine virale ne possède pas de domaine
extraceilulaire et donc e k n'a pas de ligand connu ( G i n s et al., 1997). Par conséquent,
l'oligomérisation et la transmission de signal sont des propriCt6s constitutives chez LMP-1 lui
permettant d'activer Les cellules et de favoriserla persistance de 1'~ectionvirale.
Figure 14. Modèle théorique de la cascade de ~gnalisationcellulaire déclenchée par LMP-1.
D'aprh Hiopoulos et al., 1998.
La molécule est wnstituée de t r a s domaines principaux (voir figure 13) :
Une courte queue cytoplasmique NH2-tenni.de.
Un domaine membranaire comportant six portions trans-membranaires, responsable de
l'oligomérisation spontanée de la molécule.
Enfiin, un deuxième domaine cytoplasmique, la queue COOH-terminale, longue d'environ
20 acides aminés, responsable de l a transmission de signai. La partie proximale de cette
queue, appelte CI'ARl (pour région activatrice carboxy-terminale), est capable de lier l u
protbines TRAF1, TRAF3. T W 5 et a un moindn degré TRAF2. Cette voie de
signalisation est responsable A 25 % de I'activation du facteur d e transcription NF-KB,
probablement a la suite d'une interaction avec la p d i n e NIK comme c'est le cas chez
CD40 (voir figure 5, page 26). La partie distale de la queue cytoplasmique, ou CïAR2,
est également capable de recruter T U F 2 par l'intermédiaire de la protéine TRADD
(domaine de mort associé au récepteur du TNF). Cette deuxième voie de signalisaîion est
responsable de l'activation de NF-KB à 75 46. Il semble que ces deux voies rCgulent
1'expression de g h e s differents.
A priori, puisque LMP-1 provoque la surexpression de
CD40, on pourrait croire que
l'engagement du récepteur par CD154 résulterait en une ltponse (prolif6ration) accrue chez
les lymphocytes B porteurs du virus. Cependant, aucune action synergique n'a Cté rapportCe
Lors de la stimulation de LCL via CD40 (Kilger et al.. 199û). Au contraire. la Litterature fait
même mention d'une inhibition de la prolifkration (Bishop et al., lm).De même, chez des
souris SCID, où le transfert de cellules porteuses d'EBV cause habituellement le
développement de tumeurs reliées au virus. le traitement avec un anti-CD40 a pour effet de
limiter la proliferation ceiiulaire (Murphy et al., 1995).
Plusieurs facteurs peuvent expliquer l'apparente " i m m ~ t é "des lymphocytes B A la
stimulation via C M . La premi6re hypothèse repose sur le fait que l'inftction d'un
lymphocyte B par EEW cause non seulement la régulation positive de l'expression de C D 4 ,
mais egalement la sécr6tion de cette molCcule sous une forme soluble. De plus cette forme
soluble de CD40 (ou CWOs) s'av2re capable de fuer ligand CD154 exprim6 La surface des
lymphocytes T. Les cellules des patientes atteintes de LPCB, dont une proportion est infectée
par EBV, pourraient ainsi relâcher du 0s.Or le système de culture employé dans ce projet
pour cmctCriser les lymphocytes B est basé sur l'engagement de la molCcule CWO exprimée
à la surface de ces derniers par son ligand exprimC par les œliules adhérentes LAS.
Dans
l'éventualité oh la molécule CD40s serait sécrétée dans le milieu de cultare, eue ailrait donc
pour conséquence d'entrer en compétition pour L'engagement de CD154 et CvcntueUement de
bloquer compléternent l'accès du ligand aux lymphocytes B. il est plausible d'affumer,
surtout lors de cultures B long terme. que ce ph6nomène ait Cté la cause de l'absence de
prolif6raâion observée lors de la stimulation des ailules de patientes. En outre, si la
production de CD-
est fluctuante, cela pourrait élucider les difEcultés éprouvées B
reproduire les résultats obtenus lors de la stimulation de U3L dans le système de culture
ddpendant de CD4û-
Une autre hypoth8se se base plutôt sur les inted6rences pouvant exister entre les mol€cules
CD40 et LMP-1. Comme mentionnk précédemment, les deux protéines ont des propriétés
fonctionnelles analogues :fmt les mêmes molécules, déclenchant des voies de signalisation
semblables et aboutissant ii l'activation d'un même facteur de transcription (NF-KB), elles
mènent à l'activation et la prolifhtion cellulaire. Kilger et collaborateurs (1998) ont même
demontré que, chez des LCL infectbes par un virus exprimant conditionellement la protCine
LMP-1 sous le contrôle de la tttracycline, l'activation via CWO peut complètement suppléer B
l'absence de LMP-1. Il semble donc que les deux molécules soient interchangeables.
Néanmoins, pour certains, c'est cette homologie existant entre les voies de signalisation de
CD40 et LMP-1 qui empêcherait toute action synergique.
En effet, la concentration
cytosoiique et la disponibilit6 des molécules TRAFZ et TRAF3 paraissent moduler la réponse
à la stimulation via CWO chez les lymphocytes B (Kuhné et d.,
1997). Or, le recrutement de
TRAFZ et TRAF3 par LMP-1 diminuerait la disponibilit6 de ces mol6cules pour lier
subséquemment CD40, provoquant l'inhibition de la signalisation via ce dernier recepteur. On
ferait face à un phénomène de désensibilisation (Wunt5 et ai., 1997) qui expliquerait l'absence
de prolifikation en réponse it CD40 observbe chez les LCL ou chez cellules de patientes
porteuses du virus. Toutefois, il reste B élucider pourquoi la proportion des cellules de
patientes qui ne sont pas porteuses du v h s ne prolifèrent pas même après 2û joua de culture.
Peut-être fait-on face aux effets combin6s de la sécr€tionde Os,Moquant l'accès du ligand
CDlW aux lymphocytes B pouvant théoriquement répondre A la stimulation, et A la
ddsensibilisationdes autres cellules porteuses du virus.
60
Si plusieurs études font mention de l'inhibition p16~uméede la croissana ceiiulatirc
exercée par CD40 sur les alIulcs infcctdes par EBV, peu d'entres-elles ont pu d6temher les
mécanismes précis qui en d e n t la cause. Cependant, les résultats obtmus lors de la culture
d'une LCL dans l e système de culture dCpenâant de l'activation par CWO permettent de
proposer un m€canisme expliquant ce phénomène. Ainsi, 5jours a p r b la mise en culture. une
forte baisse de la pmlif6ration cellulaire a €te constatee. Des analyses &ectu&s
par tests
TUNELont indique que cette baisse Ctaït accompagnée une hausse notable de I'apoptose chez
les cellules en culture. Ce phénomène d'induction de l'apoptose par CWO a déjà Cté rapport6
dans la Iittérahire chez des cellules c a n d p s a d'origine épithéliales et mésenchymiales
(Hess et al.. 1996), mais les m€can.ïsmes qui le dCclcnchent demeurent inconnus ik ce jour. L+
système de stimulation in vin0 par CWO ainsi que la disponibüté de plusieurs LCL offrent
l'occasion d'approfondir 1'Ctude des conséquences de la stimulation via le récepteur chez des
cellules porteuses du virus EBV. Auparavant, il sera toutefois nécessaire d'améliorer certaines
conditions de culture. Il faudra notamment s'assurer que le ligand CD154 est toujours présent
en excès, ce qui poumit être achev6 en utilisant une forme trim6risée soluble de la molécule.
De plus le test TUNEL ayant certaines limites (particulièrement en ce qui à hait B la
distinction entre la nécrose et l'apoptose), il sera bon d'utiliser une seconde methode de
détection de l'apoptose pour conflmer les résultats obtenus. A ce titre l'anticorps M30 mis
sur le marché récer~mentpar Boheringer Mannheim et qui reconnaît spécifiquement les
cellules apoptobiques paraît être une b 0 ~ option.
e
Ces précautions devraient permettre la
reproduction des expériences et assurer la fiabilité des résultats.
CONCLUSION
La lymphocytose polyclonale chronique B est un désordre du systhne hématologique qui
n'a Cte ddcouvert que récemment et qui demande encore B être catactérisC. N o m projet
explorait un aspect de la maladiejamais abord6 encore. soit la caractérisation fonctionnelle des
lymphocytes B et T de patients, dans le dessein de complCter les ciifferentes Ctudes
morphologiques et g6n6tiques publiCes jusqu'8 présent. Les r&uitats obtenus ont démontré
sans équivoque que les cellules des patients se distinguaient remarquablement des cellules de
témoins normaux par leur incapacitC à répondre au signal de prolirdration et d'activation
cellulaire engendr6 par la molCcule C M . Or il se trouve que cette molécule joue un rôle
prépondérant dans le cycle cellulaire des lymphocytes B, le développement de la réponse
immunitaire, le maintien de l'hom6ostasie du système immunitaire. En outre, les données
expérirneniales appuient I'hypothhe selon laquelle Le virus Epstein-Barr jouerait un rôle dans
la maladie. Toutefois, de nombreuses interrogations sont aussi soulev6es. Ainsi, il n'est pas
encore possible de faire la discrimination entre cellules normales et cellules anormales chez
les patients. Est-ce que toutes les cellules possMent un defaut intrinshpe interf6rant avec la
stimulation via CWO ? Ou est-ce qu'alternativement seules les ceiiuies porteuses du virus
sont affectées ? Ces questions soulignent l'importance de trouver un marqueur permettant
d'identifier spécifiquement les cellules qui seraient à la source de la lymphoprolif'ration.
D'un autre côté il n'est pas possible A ce point d'Ctablir quoi, de l'infection virale ou du dtfaut
fonctionnel, serait & l'origine de la LPCB. Est-ceque la rCponse la stimulation via CD40 est
perturbée en conséquence de l'infection virale des lymphocytes B ? Ou bien est-ce la
désensibilisation aux effets induit par le rkeptew qui permettrait la survie des cellules
infectées devant normalement être élimin6es ? L'expression accrue des ghes de latence du
virus chez les patients laisse en effet planer des douies sur 1' aptitude du syst8me immunitaire
il contrôler efficacement l'infection virale.
En conclusion bien que les résultats ce projet apportent de nouveaux Cltments & la
caractérisation de la LPCB,il reste beaucoup de points B Cclakir pour en cerner l'origine du
syndrome et comprendre les mécanismes menant au développement de la lymphocytose.
BIBLIOGRAPHIQUES
Agrawal, S. Matutes, E,Voke, J, Dyer, MIS, Khokhar, T,Catovsky, D. (1994) Persistent
B-Ce11 Lymphocytosis. Leuk.. Res. 18: 791-795.
AUday, MJ, Crawford, DH. ( l m ) Role of Epithelium in EBV Persistena and
Pathogenesis of B-CeIl Turnours. -et
1: 8 5 M -
Aimarri, A, Batchelor, IR ( l m ) HLA and Hepatitis B Mixtion. Lmicet 344: 11%
1195.
Baer, R, Bankier, AT, Biggin, MD, Deininger, PL, F d ,PI, Gibson, TJ, Hatfull, G,
Hudson, GS, Satchwell, SC, Seguin, C. (1W)DNA Sapence and Expression of the
B95-ûEpstein-Barr Virus Genome. N-e
310: 207-21 1.
Banchereau, J, De Paoli, P, Vallt, A, Garcia, E,Rousset, F. (1991) Long-term Human B
Celi Lines Dependent on Interleukul-4 and Ancibody to CD4û.. Science 251: 71-72.
Banchereau, I, Bazan, F, Blanchard, D,Brière, F, Galizzi, JP, Van Kooten, C, Liu, YJ,
Rousset, F, Saeland, S. (1994) The CD40 Antigen and Its Ligand. Annu. Rev. I m m u ~ L .
12:881-922.
Bishop, JY, Schattner, EJ, Friedman, SM.
(1998) CWO Ligation Impedes
Lymphoblastoid B Cell Proliferation and S-Phase Entry. Leuk. Res. 22: 319-127.
Cailet-Bauchu. E,Renard, N, Gauo, S, Poncef C, Morel, D, Pagès, J, Salles, G, Cœur,P.
Felman, P. ( 1997) Distribution of the Cytogenetic Abnormality +(3)(q10) in Persistent
Polyclonal B-CeU Lymphocytosis: a FICIION study in thme cases. Br. J. Haemcltol. 99:
53 1-536.
Cam, R, Fishlock, K. Matutes, E. (1997) Persistent Polydonal B-CeII Lymphocytosis in
Identical Twins. Br. J. Haematol. 96r 272-274.
Cars&,
KC, Fmcombe, WH, Scott, JG, GeIfand, EW. (1985) Persistent Polyclonal
Lymphocytosis of B Lymphocytes, Induced by Cigarette Smoking? Luncet 1: 10W.
Chow, KC, Naciiia, JQ, Witzig, TE, Li, CY. (1992) 1s Persistent Polyclonal B
Lymphocytosis Cawd by Epptein-Barr Virus? A study with Polymerase Chain Reaction
and in Situ Hybridation. Am. I. Hemutol. 41: 270275.
Delage, R, Darveau, A, Jaques, L, Huot, A, Delage.
JM. (1992) Chtonic B Ceîl
Lymphocytosis of the Young Woman: Clinid, Phenotypic and Molecular Studies in 5
Cases. B W 80 supp.: 4 7 a .
Delage, R Roy, J, Jacques, L, Bernier, V, Delage, JM, Darveau, A. (1997) Multiple
b c M g Gene Rearrangements in Persistent Polyclonal B-Ceil lymphocytosis.
Br. J-
Haematol. 97: 589-595-
Delage, Ft, Roy, J, Jacques, L, Darveau, A. (1998) AU Patients with Persistent Polyclonal
B-CellLymphocytosis Present Bcl-2flg Gene Ruurangments. Leuk. Lymph. sous presse.
De-Thé,G. (1982) Epidemiology of Epstein-Barr V i s and Associated Diseases in Man.
Dans nie Hevemiruses. Roimüum B (editeur), New-Yorkand London, Plenum h s s ,
vol. 1, p. 151-205.
Devergne, O, McFarland, EC, Mosialos, G, Izumi, KM, Ware, CF, Kieff, E. (19%) Role
of the TRAF Binding Site and NF-KB Activation in Epstein-Barr Virus Latent Protein 1Induced Ce11 Gene Expression. J. Virol. 72: 7900-7908.
Dewhurst, S. (page consultée le 28 aofit 1998) Herpesvirus: Lecture 2 (en ligne). Adresse
URL: h~p://www.urmc.r0~he~ter.edu/~mdmbi/grad/he~p97B.html.
Duchosal, MA. (1997)B-Cell Development and Differentiation. Seminms Hemufol. 34
supp.: 2- 12.
Eliopoulos, AG, Ricichson, A. (1998) Epstein-Barr Vi: LMPl Masquerades as an
Active Receptor. C m .Biol. 8: R196R198.
Epstein, MA, Achong, BG, Barr, YM. (1%4) Virus Particles in Cultured Lymphoblasts
from Burkitt's Lymphoma. Lancez 1: 702-703.
Faris, M,Gaskin, F, Parsons, JT,Fu, SM. (1994) CD4û Signalhg Pathway: Anti-CD40
Monoclonal Antibody hduces Rapid Dephosphorylation and Phosphorylation of TyrosinePhosphorylated Roteins hcluding Rotein Tyrosine Kinase Lyn,Fyn, and Syk and the
Appearance of a 2gkD Tyrosine Phosphorylated Protein. J. Exp. Med. 179: 1923-1931.
Farrell, PI. (1995) Epstein-Barr Virus Irnmortalizing Genes. Trends Microbwl. 3: 105-
109-
U,Gonneila, R Uentng, M. Marschall, G, Zeidler, R, Pich, D,
Hammerschmidt, W. (1991) Latent Membrane Protein 1 of Epstein-Barr V m Mimics a
G i n s , O, Zimber-Strobl,
ConstitutivelyActive Reœptor Molecule. EMBO 16: 6131-6140.
TJ,Lawrcna, DN. (19â2) Persistent
Polyclonal Lymphocytosis of B Lymphocytes. Nov EngL J. Med. 3û7: 233-235.
Gregory, CD, Dive, C, Henderson, S. Smith, CA. Williams, GT, Gordon, I, Rickinson.
Gordon, DS, Jones, BM, Browning, SW, Spira,
AB. (1991)Activation of Epstein-Barr Virus Latent Genes Rotects Human B Ceils h m
Death by Apoptosis. Nmure 349: 612614.
Hanissian, SH, Geha, RS- (1997) JAK3 1s Associateci with CD40 and 1s Critical for CD40
Induction of Gene Expression in B Ceus. Imm-ty
Hanto, DW.
6: 379-381.
(1995) Ciassification of Epstein-Barr Virus Associaîed Posttransplant
Lymphoproliferative Diseases. AMII. Rev. Med. 16: 381394.
Heath, AW, Chang, R, Hatada, N, Santos-Argumedo, L, Gordon, J, Hannum, C,
Campbell, D, Shanafelt, AB, Clark, EA,Toms, R Howard, M. (1993) Antibodies to
Murine CD40 Stimulate Normal B Lymphocytes but Inhibits Roliferation of B
Lymphoma Cells. CefZ Immunul. 152: 468480.
Hess, S, Engelmann, S. (1996) A Novel Function of CWO: Induction of CeU Death in
Transfonned Cells. J. Exp. M d 183: 159-167.
Hsing, Y, Hostager, BS, Bishop, GA.
(1997) Cbaracterization of CWO Signaling
Detemïnants Regulating Nuclear Factor-KB Activation in B Lymphocytes. J. I m n o l .
159: 489û-4906,
Karajamis, MRT Huemmei. M, Anagnostopoulos, 1, Stein, H.
(1997) Strict
Lymphotropism of Epstein-Barr Vis During Acute Infectious Mononucleosis in
Nonimmunocompromised Individuals. Bbod 89: 2856-2862.
Kehry, MR. (19%) CD40 Mediated Signahg in B-celi. J. Inununol. 156: 2345-2348Khanna, RTBurrows, SR, Moss, DJ. (1995) Immune Regulation in Epstein-Barr Virus-
Associated Diseases. Microbiol. Rev. 59: 38745.
Khun6, MR,Robbiis, M, Harnbor, JE, Mackey. MF, Kosaka, Y, Nigishimura, TTGigley,
JP,Noeile, RI, Calderhead, DM. (1997) Assembly and Regulation of the CD40 Receptor
Complex in Huma,B Ceils. J. Exp. Med. 118L: 337-342.
Kieff, E,Liebowitz, D. (1990)Epstein-Barr V i and itr Repiication. Dans Virology.
ed. Fields
BN, Knipe DM,HowIey PM (éditeurs), New-York, Raven Press, p.
1
-
1920,
Kilger, E, Kieser, A, Ba-,
M, Hammerschmidt, W. (1998) Epstein-Barr Virus-
Mediated B-CeU ProIifkration is Dependent upon Latent Membrane Protein 1, which
Simulates an Adivaîed CD40 Receptor. EMBO 17: 170-1709.
Lapointe, R Lemieux, R, Olivier, M, Danieau, A. (19%) Tyrosine Kinase and CAMPDependent Rotein Kinase Activities in CD40-Activated Human B Lymphocytes. Ela. J.
Iktmunol- 2& 2376-2382,
Larcher, C, Fend, F, Mitterer, M, Rang, N, Schwamnann, F, Huemer, HP. (1995) Role of
Epstein-Zarr Virus and Soluble CD21 in Persistent Polyclonal B-CellLymphocytosis. Br.
J. Hàema.tol. 90: 532-510Law, C , Craxton, A, Otibopy, KL, Sidorenko, SP, Klaus, SI, Clark, EC. (1996)
Regulation of Signaling through B-Lymphocyte Antigen Receptors by Cell-Cell
Interaction Molecuies. Imunol* Rev. 153: 123- 154.
Malinin, NL,Boldin, MP,Kovalenko, AV, Waliach, D. (1997) MAP3K-Related Kinase
Involved in NF-KBInduction by TNF, CD95 and L I . N m e 3û5: 5QO-544-
Marelie, L, RCa, D, Raphaël, M. (1993) Le virus d'Epstein-Barr et les prolX6rations
lymphoïdes. Med.Sci- 9: 693-700.
C, Rang, N, Schwamnann, F, Coser, P,
Huemer, HP. (1995) Chronic Active Epstein-Barr Virus Disease in a Case of Persistent
Mitterer, M, Pescota, N, Fend, F, Larcher,
'
Polyclonal B-Celi Lymphocytosis. Br. Hczematol. 90: 526-53 1.
Murphy, WJ,Fimakoshi, S. Beckwith, M. Rushing, SE, Conley, DK, Armitage, RJ,
Fanslow, WC, Rager, HC, Taub, DD, Ruscetîi, FW, Longo, DL. (1995) Antibodies to
CD40 Revent Epstein-Barr Virus-Mediated Human B CeU Lymphomagenesis in Severe
Combined Immune Deficient Mice Given Human Peripheral Blood Lymphocytes. Blood
86: 194tG1953.
Nkroo, S, Pelletier, A, Chevrier,M, Monier, G, Lemieux, R, Darveau, A- (1996) induction
of LAF-1 Independent Human B Cell RoiXeration and DZferentiation by Binding of
CD40 with Its Ligand. Immunal. Imest. S.79-89.
Niedobitek, G, Young,LS. (1994) Epstein-Basr Virus Penistencc and ViAssociatcd
Turnours. Lance? 343: 333-335.
Pelloquin, F, Lamelin, JP, Lenoir, GM. (19%)
Human B Lympbocytes Immortalization
by Epstein-Barr Vinis in the Presence of C y c l o s p o ~
A. In Viao CeIl. & Devel. Biol. 22:
689-693Perreault, C , Boileau, J, Gyger. M., De BeiideuilIe. C, DTAngelo, G, Bélanger. R,
Lacombe, M, Lavallée, R, Bomy, Y, Paquin. M. Bmchu, S.
(1989) Chronic B-CeU
Lymphocytosis. Ela. J. Haematol. 42: 361-367.
Rickinson, AB, Moss, DJ. (1997)Human Cytotoxic T Lymphocyte Rcsponses to Epstein-
Barr Virus Infwtion. Annu*Rat* Immunol, 15: 405-43 1.
Robinson, JE, Miller, G. (1982) Biology of Lymphoid Celis Trandormed by Epstein-Barr
V h s . Dans The Herpemiwes. Roiunam B (éditeur), New-York and London, Plenum
Press, vol. 1, p. 151-205.
Rocchi, G, De Fellici, A, Ragona. G, Heinz, A. (197ï)Quantitative Evaluation of Epstein-
Barr Virus-Infected Mononuclear Blood Leukocytes in Infectious Mononucleosis. N.
Engl. J. Med. 296: 132-134.
Rothe, M, Sarnia, V, Dixit, VM, Gaddel, DV. (1995) ïïWF2-Mediated activation of
NF-KB by TNF Receptor 2 and CD40. Science 269: 14%-142%
Roy, J, Ryckmann, C, Bernier, V, Whittom, R, Delage, R. (1998) Large CeU Lymphoma
Complicating Persistent Polyclonal B-Cà1Lymphocytosis. Leukemia 12: 1026lm.
Ryckman, C,Darveau, A, Delage, R (lm)Molecular Analysis of Epstein-Barr Virus
Gene Etpression in Persistent Polyclonal B Ceil Lymphocytosis. En soumission.
Thorley-Lawson, DA, Miyashita, E, Khan, G. (1996) Epstein-Barr Virus and the B CeU:
That's AU it Takes. Trends Microbiol. 4: 2W2û8.
Tosato, G, Kazuyuki, T, AngiolUo, AL, Sgadari, C. (1995) Epstein-Barr Vinis as an
Agent of Haematological Disease. Builli&e 's Clin. Hàemztol. 8: 161186.
Troussard, X,Valensi, F, Debert, C, Maynadie, M, Schillinger, F, Bonnet, P,Macintyre,
EA, Flandrin, G.
(1994) Persistent Polyclonal Lymphocytosis with Binucleated B
Lymphocytes: a Genetic Predisposition. Br. J. Haematol. 88: 275280.
Troussard, X. Mossafa, H, Valensi. F,Maynadie, M,Schillinger. F. Bulliard. G.Malamt
H,Flandrin. G. (1997a) Lymphocytose polyclonale avec lymphocytes sanguins binucléés.
La Presse Mi5dicczle 2 6 895-899.
Troussard, X. Mossafa, HPRancirin, G. (lm)Identity between Hajr B-Cell
Lymphoproliferative Disorder and Persistent Polyclonal B Lymphocytosis? B M 9û:
2110-2113Van Kooten, C,Banchereau, J. (1997) Immune Regdation by CWO-CWOL Interactions.
Fronziers in Bioscience. 2: 1-11.
Wfiams, M.(1990) Lymphocytosis. Dans Hematology. 4* cd. Williams, WJ,Beutler,
E, Erslev, AJ,Litchman, MA (éditeurs), New-York, McGraw Hill inc., p. %3-%4.
Young, LS, Sixbey, W. Clark, D,R i c h o n , AB. (1986) Epstein-Barr V i Receptors
on Human Pharyngeal Epithelia k e t 1: 240-242Zubler, RH. (1997) Key Differentiation Steps in Normal B Cell and in Myeloma Cells.
Serninars Wematol. 34 supp.: 13-22.
ANNEXE A.
Optimisation des conditions de cdtpro poar la Samadation mitgémique par h PHA.
FigoR 15. Optimisation du test de stimulation ii la PHA en fonction de la concentration
celIulaire. Des concentrations de PBMC variant de 2.5X10' B lXlo6 celluIedpuits ont et6
cultivées en triplkata en prCsence de PHA (valeur optimale thhique de 1.5 pghl), dans une
plaque de % puits B fonds plats, 48 heures B 37T, 5 8 CO,. La prolif6ration cellulaire a alors
été évaluée par test colonm&ique MIT (Iecture B 550nm,diCrence de 690 nm).
F
i 16. Optimisation du test de stimulation à la PHA en fonction de la concentration de
mitogène. .%le
PBMUpuits ont €te cultivées en présence de PHA & une concentration finale
de 0.5, 1.25,2.5,3.75 et 5 pg/ml. Les cultures ont C t é effectuées en triplicata dans une plaque de
% puits à fonds plats, 48 heures A 3TC, 5 96 CO,. La proIifCration cellulaire a alors €te 6valuCe
par test colorim€triqueMTT' (lecture B 550nm, reference de 690nm).
incubaam (h)
Figure 17. Cinétique du test de stimulation B la PHA. 5x10'PBMUpuits ont Cté cultivées en
triplicata dans une plaque de % puits à fonds plats, en prbsence de PHA à une concentration
finale 1 pg/rnl. Les cultures ont ét6 effectuées sur des périodes de 48, 72,% et 120 heures à
37OC,5 % CO,. La prolif€ration cellulaire a alors Cté Cvalude par test c o l o ~ é û i q u e
(lecture à 550nm,rbference de 690nm).
En conclusion, la stimulation par la PHA sera optimale dans les conditions de culture
suivantes :
Une concentration cellulaire de 5x105celluledpuits.
6 Une concentration de PHA de 1 pg/mi.
7 Une durée d'incubation de 48 heures.
5
ANNEXEB
Détermination de Ia proportion âe lympocytes T chez la PBMC de paüeates.
Tableaa 5 :C a i c d de ia concentration c e n m en fonction da nombm de ipnphocytes
Tlpaig: La proportion de lymphocytesT des diff6rcntcs populations de PBMC utilisées dans
le test de stimulationPHA a Cîé Cvaluée par cytofluoromdtrie. Les pourcentages obtenus ont
6té utiiisés pour convertir la concentration ceUulain exprimée en ceiiuiesfpuits en
lymphocytes T/puits.
1
ceIldes /puits
1 TCmoins
I
(7096 IymT)
1 Patiente B
1 Patiente M 1 Patiente S
(41.5 46 IymT)
(34.2% IymT) (34.6 % lymT)
855 XI@
8.65x l e
1.75~
le
1.~14 x l d
5xld
2.5 XI@
2.08 x i d
1.71 x l d
1.73 x l d
7.5xld
5.25 xi05
3.1 1 xL05
2.57 xi@
2.6xld
1x10~
7x l d
4-15 xld
3.42 x l d
3.46 xld
1.25x106
8.75 x l d
5.19 xlv
4.28 x l d
433 x l d
2 . 51
~d
I
I
ANNEXE C.
Stratégie d96valuationde I'immdté dulaire spécif@ne
IEBV.
1- Génération de LCL
Lymphocyte B
EBV
LCL
2- Génération de CTL spécifiques à EBV.
Lymphocyte T
LCL
irradik
3- Mesure de la cytotoxicité spécifique à EBV.
O
:::::::::
:::::::::::::
:::::::::::::
.-..a-.-...-.
-.-.-.-
::::y::::
LDH
-
P-
LDH
(absorbe à 500 nm)