Exercices Corrigés Système Nerveux: Biologie

1
CORRECTION EXERCICES sur le SYSTEME NERVEUX
I) RESTITUTION DES CONNAISSANCES
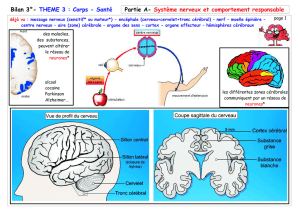
1) Schémas à titrer et à légender
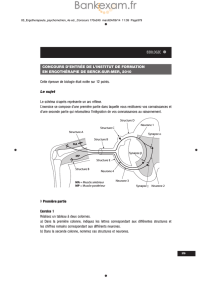
Schéma 1 :
Titre : Coupe Transversale de la colonne vertébrale et de la moëlle épinière dans la région
cervicale
1. Cloison névroglique
2. Corne postérieure ou dorsale
3. Arachnoïde
4. Pie Mère
5. Substance Blanche ou matière blanche
6. Canal de l’épendyme
7. Sillon antérieur ou ventral
8. Corne antérieure ou ventrale
9. Racine antérieure ou ventrale
10. Racine postérieure ou dorsale
11. Ganglion spinal ou rachidien
12. Dure-mère
13. Tissu adipeux
14. Vertèbre
Schéma 2 :
Titre : Coupe Transversale montrant quelques éléments d’un arc réflexe polysynaptique.
a. Racine dorsale du nerf rachidien
b. Péricaryon du neurone en T
c. Ganglion rachidien (= spinal)
d. Motoneurone a de l’extenseur
e. Racine ventrale du nerf rachidien
f. Motoneurone a du fléchisseur
g. Corne ventrale (substance grise)
h. Cordon latéral (substance blanche)
i. Corne dorsale (substance grise)
j. Interneurone inhibiteur
2) Questions issues de différents concours
a) La longueur du muscle.
b) Le neurone est l’unité structurale et fonctionnelle du tissu nerveux. Le neurone
possède des propriétés d’excitabilité et de conductibilité. Le neurone ne se multiplie
pas.
c) Le récepteur sensible à un étirement léger du muscle est le fuseau neuro-musculaire.
d) Formation d’un potentiel générateur à l’origine d’un PA qui gagne alors le centre
nerveux en empruntant le prolongement central du neurone sensitif.
e) Perte de la motricité des éléments musculaires réliés au nerf rachidien concerné, la
sensibilité de cette région reste intacte.
f) Nerf composé de fibres sensitives (afférentes) et de fibres motrices (efférentes) ex : le
nerf rachidien.

2
4) QCM
1. B, E
2. E, F
3. B, D, E
4. B, D, E, F
5. B, E
6. C, E
7. D, F
8. C, D, E
9. B, C, E, F
10. A, C, E
11. B, C, D, E
12. H
13. E
14. B
15. Aucune réponse inexacte
16. A – G – E – F – B – D – C – H
17. A, C, D
18. B, C
19. B, D, E
20. B
21. D
22. A
23. B
24. C
25. C
II) EXERCICES
Exercice 1 :
Neurotransmetteur Hormone
Sécrété par Un neurone Une cellule endocrine
Agit * Sur son lieu de sécrétion,
ne parcout que quelques mm.
* Atteint rapidement sa cible
et agit donc rapidement.
* Action très localisée
* A distance, par diffusion et
transport dans le milieu
intérieur.
* Atteint plus lentement sa
cible, agit donc plus
lentement.
* Action diffuse
Mode d’action Par l’intermédiaire de
récepteurs spécifiques Par l’intermédiaire de
récepteurs spécifiques
Dégradation Rapide (quelques ms) dans la
fente synaptique Plus lente (quelques min ou
quelques jeures) dans le
milieu intérieur
Effets Modifie l’état électrique de
l’élément post-synaptique Modifie le métabolisme,
l’action d’enzymes …
Exemples Ach, GABA … Insuline, Glucagon ….
Le système hormonal et le système nerveux sont 2 systèmes de communication de l’organisme. Ils
participent à l’unité physiologique de l’organisme car ils mettent en relation les organes les uns aux autres
et assurent la cohésion fonctionnelle de ces organes.
Dans les 2 cas, des messagers chimiques (neurotransmetteurs et hormones) interviennent, ceux-ci sont
libérés par des émetteurs de messages (neurones ou cellules endocrines) et agissent sur des cellules cibles
spécifiques (via des récepteurs).
Dans le cas du système nerveux, la majorité du trajet entre l’émetteur du message et la cible est assurée par
des PA, propagés à vitesse importante.
Dans le cas du système hormonal, la plus grande partie du trajet entre émetteur de messages et cible est
assurée par une mise en circulation de l’hormone. Celle-ci atteint plus lentement sa cible.

3
Dans tous les cas, le codage de l’information est assurée par une modulation de la concentration du
messager.
Exercice 2 :
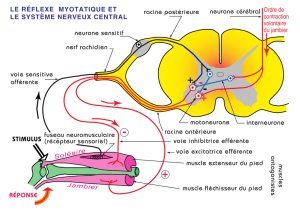
La percussion effectuée sous la rotule provoque un étirement du muscle extenseur de la jambe. En réponse,
il se produit une extension de la jambe, c'est-à-dire d’une contraction du muscle extenseur qui a été étiré. On
est en présence d’un réflexe myotatique. Cependant, il faut coordonner cette réponse avec un relâchement
du muscle antagoniste c'est-à-dire le muscle fléchisseur de la jambe. Le point de départ sont les F.N.M.
Ceux-ci sont stimulés par l’étirement du muscle extenseur
Ils émettent des signaux nerveux en direction de la moelle épinière (contre intégrateur) via des neurones
sensitifs. Dans la substance grise médullaire le système diverge.
D’une part, il y a une synapse excitatrice entre la voie sensitive et le motoneurone du muscle qui a été étiré.
Ce muscle va alors se contracter.
D’autre part, il y a une synapse excitatrice avec un interneurone qui, à son tour, va inhiber le motoneurone
du muscle antagoniste (muscle fléchisseur) lequel va se relâcher.
L’action coordonnée sur les 2 antagonistes permet une extension de la jambe. On parle d’innervation
réciproque.
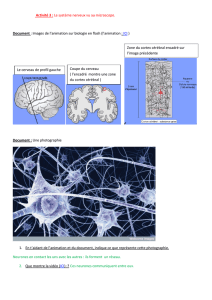
Exercice 3 :
1. L’encéphale commandant le fonctionnement de la moelle épinière, on est certain, en
déconnectant les 2 centres nerveux, que les réponses obtenues sont seulement dues aux
stimulations délivrées dans l’expérience.
2. La stimulation du bout de la fibre Ia relié à la moelle épinière entraîne une réponse du muscle.
Cette fibre véhicule des informations vers la moelle épinière. Seule la stimulation du bout
périphérique de la fibre a entraîne une réponse. Les informations véhiculées par cette fibre
circulent en direction du muscle. On peut établir le sens de circulation du message nerveux :
Fibre Ia => ME => motoneurone et fibre a

4
3. La fibre Ia est une fibre sensitive.
La fibre a est une fibre motrice.
Exercice 4 :
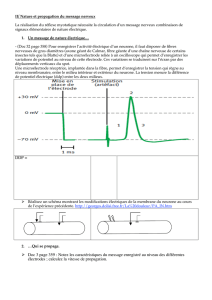
A) Avant la pénétration de la micro-électrode R1 dans l’axone, le potentiel mésuré en mV par
l’oscilloscope est nul.
Lors de sa pnétration, ce potentiel chute à -60 mV, ce qui montre qu’il existe une ddp de part et d’autre
de la membrane axonique. Cela montre que la membrane est POLARISEE : la face externe étant
chargée positivement et la face interne négativement.
Cette ddp de -60 mV représente le potentiel de repos ou potentiel transmembranaire.
B) Le premier tracé est un artéfact de stimulation, il n’est pas suivi d’effet, l’intensité de la stimulation a
été insuffisante. On parle d’intensité infraliminaire.
En revanche, les deux stimulations suivantes sont efficaces, elles sont suivies avec un temps de latence
d’une dépolarisation dont le pic atteint +60 mV (b et c). Le pic de 60 mV est le PA.
Puis la ddp revient à sa valeur initiale (potentiel de repos) après une période d’hyperpolarisation.
On remarque qu’en b et c la variation de potentiel est identique l’amplitude du PA est indépendante de
l’intensité de la stimulation. Sur un neurone, selon son intensité, une excitation provoque ou non la
naissance d’un PA : c’est la loi du tout ou rien.
Mouvements ioniques (revoir le cours)
Exercice 5 :
1. L’analyse des tracés obtenus suite à l’application de stimulations d’intensité croissante permet de
mettre en évidence :

5
- la présence d’un seuil d’intensité pour l’apparition d’une réponse. En dessous de ce seuil
(stimulation S1), aucune réponse n’apparaît : on parle de stimulation infraliminaire. Au dessus de ce
seuil, un PA est généré, on parle de stimulation supraliminaire. Ce PA est caractérisé par une phase
de dépolarisation suivie d’une phase de repolarisation.
- le fait que la réponse de la fibre n’est pas graduable. Lorsque le PA apparaît, il a toujours la même
amplitude quelle que soit l’intensité de la stimulation.
La fibre nerveuse répond selon la loi du tout ou rien. La réponse est non décrémentielle.
2. Analysons les réponses obtenues en éloignant les électrodes stimulatrices et réceptrices. On constate
que les points A1 et A4 sont alignés et définissent une droite de pente constante. Cette pente
correspond à la vitesse de propagation des P (v = d / t = Y / x).
La vitesse est donc constante sur toute la longueur de la fibre. La pente de la droite est :
Y4 – Y1 / x4 – x1 = 35 – 10 / 1,03 – 0,34 = 36
Donc v = 36 mm/ms soit 36 m/s.
Exercice 6 :
1. Le message nerveux moteur répond à une stimulation efficace (c'est-à-dire génératrice d’un PA)
effectuée sur les fibres afférentes. Ce n’est que dans le troisième cas (lorsqu’on stimule F1 + F2 + F3)
que ce PA va se transmettre au motoneurone M1 et se propager sur son axone (on peut l’enregistrer
en A). C’est ce PA qui constitue le message moteur qui va provoquer la contraction de fibres
musculaires innervées par M1.
Le message moteur naît si la dépolarisation dépasse le seuil critique. Celui-ci se propagera sans perdre
l’amplitude.
2. Les différents enregistrements obtenus en M1 sont les réponses à des stimulations qui concernent un
nombre croissant de fibres afférentes : F1 puis F1 + F2 puis F1 + F2 + F3. La stimulation sur la seule
fibre F1 est suivie en M1 d’une légère dépolarisation appelée PPSE (potentiel postsynaptique
excitateur). Lorsqu’on stimule F1 + F2, on enregistre encore un PPSE en M1 mais son amplitude est
plus importante : il y a eu sommation des deux réponses. Lorsqu’on stimule F1 + F2 + F3, il y a
encore sommation, mais l’amplitude de la réponse dépasse une valeur critique à partir de laquelle un
PA apparaît sur le cône initial du motoneurone M1. On appelle la propriété intégratrice cette aptitude
que présente le motoneurone M1 à réaliser la somme des PPSE dont il est le siège.
3. Le Document 2 nous montre qu’en réponse à une stimulation efficace, le motoneurone M2 présente
dans le deuxième cas (F1 + F2) puis dans le troisième cas (F1 + F2 + F3) des hyperpolarisations
d’amplitude croissante. Il s’agit de PPSI (potentiel postsynapique inhibiteur), qui manifestent,
comme les PPSE, la particularité de se sommer, à la différence qu’une hyperpolarisation de plus en
plus forte nous éloigne de la possibilité d’observer un PA en M2.
L’analyse des enregistrements montre que c’est au niveau de l’interneurone que se crée un PA en
réponse à la stimulation F1 + F2. Mais la synapse que fait celui-ci avec M2 n’est pas génératrice de
PPSE ; bien au contraire, on y relève la formation de PPSI. Il s’agit d’une synapse inhibitrice et
l’interneurone est qualifié d’inhibiteur. Ce dispositif réalise ce qu’on appelle l’innervation réciproque
c'est-à-dire que la contraction d’un muscle inhibe l’antagoniste.
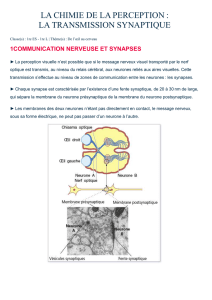
Exercice 7 : La transmission synaptique
I.
1. - Une stimulation du neurone A entraîne une réponse (activité électrique) des neurones A et C.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%