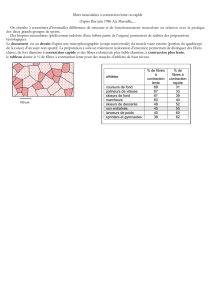
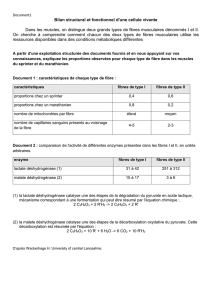
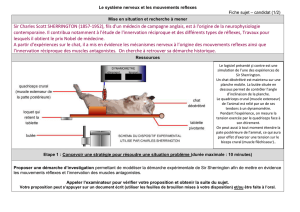
Télécharger le fichier - BP APT SXM 15-16

Le muscle et sa contraction
Comme les autres animaux, nous sommes organisés de manière à nous mouvoir, à
agir sur le milieu. La question que nous posons est : comment notre organisme se
débrouille t-il pour réaliser cette action, pour produire le mouvement ?
La réponse à cette question peut se faire à différents niveaux d'analyse. Le
premier que nous aborderons se situe à l'interface de l'organisme et de son
environnement.
1 Le regard sur le geste
Agir c'est produire une force interne à l'organisme et l'appliquer à une partie de
l'environnement. Selon les cas, cette force peut se traduire ou non par le
déplacement du corps et/ou de la partie du milieu sur laquelle elle s'applique.
Notre propos est de montrer comment cette force interne est créée et
comment elle peut se traduire par un déplacement. Voyons déjà selon quelles
grandes modalités elle peut être administrée.
1.1 Force et mouvement
Un coureur sait qu'il ne peut courir éternellement à sa vitesse maximale. Un
lanceur a bien conscience de ne pouvoir répéter x fois une grande performance à
l'exercice de développé couché. Nous savons donc que la force maximale que
nous pouvons exercer diminue avec le temps. Les scientifiques ont, eux aussi,
observé ce phénomène.
Ils ont montré que le niveau de puissance pouvant être fourni au cours
d'exercices allant de 15" à 1' connaît une décroissance logarithmique avec la
durée de l'effort. En fait, la baisse de force intervient dès les premières
secondes. Après seulement 20" d'électrostimulation sous forme 1"6 d'exercice -
1"6 de récupération, Södelund, Greenhaff et Hultman (1992) ont observé que le
muscle vaste latéral ne développait plus que 90% de sa force maximale.
Avec le temps, la force diminue. Outre le temps, des modalités purement
internes à l'organisme font varier la force pouvant être produite
1.2 Force et "état" du muscle : ce qui influence la force pouvant être
appliquée
Au moins trois caractéristiques influent sur la force générée par le muscle.
son étirement : Nous avons vu dans le secteur biomécanique (biomécanique)
que les muscles et leur tissu conjonctif peuvent, par leurs propriétés, être
comparés à des élastiques. Quand ils sont étirés, ils stockent l'énergie. Lors du
raccourcissement musculaire qui suit, cette énergie potentielle peut être

libérée.
Au repos physiologique, le muscle est très légèrement étiré. Sa longueur est
déterminée par cet étirement et par son attachement aux pièces osseuses. La
force maximale qu'il peut fournir est obtenue lorsqu'il est préalablement étiré
d'une longueur correspondant à environ 20% de sa longueur de repos. C'est pour
cet étirement que la force de contraction musculaire et la restitution de
l'énergie potentielle élastique s'additionnent de manière optimale. Au-delà ou en
deçà de ces 20%, la force développée diminue.
l'angulation : la force développée dépend de l'angle de l'articulation jouant le
rôle de levier. Par exemple, le biceps (muscle du dessus du bras) est plus fort
lorsque l'avant bras fait un angle de 90° avec le bras que lorsque cet angle est
supérieur.
la vitesse de contraction : Pour une contraction de type concentrique (quand
le muscle se "concentre"), plus le mouvement est rapide plus la force applicable
est faible. L'inverse est vrai pour une contraction excentrique.
Pour le moment, nous avons raisonné au niveau du muscle pris comme un tout
homogène. Tentons, à présent, de faire pénétrer notre regard à l'intérieur de ce
muscle. Voyons s'il peut être décomposé en parties plus petites que lui-même.
Entrons en lui pour étudier son histologie (histologie = science des tissus
constituant les êtres vivants).
2 Le regard sur les tissus
Notre regard se situe entre l'environnement et les différents tissus
musculaires.
2.1 Les différents types de fibres musculaires
Les physiologistes distinguent deux grands types de fibres. Les fibres I
appelées également fibres lentes (slow twitch - ST) ou fibres rouges et les
fibres II nommées aussi fibres rapides (fast twitch) ou fibres blanches.
Comme leur nom l'indique, les fibres I sont à contraction lente. Elles sont en
revanche peu fatigables. De faible diamètre, elles bénéficient d'une bonne
irrigation sanguine ce qui explique leur autre nom de fibres rouges. Comprenant
de nombreuses mitochondries, elles privilégient la filière aérobie.
Les fibres II sont à contraction rapide. Elles se "fatiguent" rapidement. De gros
diamètre, elles sont peu irriguées mais disposent d'importantes réserves de
glycogène. Elles privilégient la filière anaérobie.
Autre différence, plus la fibre est rapide, plus elle renferme un réticulum
sarcoplasmique développé. Que sa quo un réticulum sarcoplasmique ? Un
ensemble de sacs liés entre eux et qui gardent le calcium nécessaire à la
contraction de la fibre musculaire. Une fibre rapide est ainsi plus apte à libérer
le calcium dans le muscle donc à se contracter rapidement. Nous comprendrons
mieux le rôle du réticulum lorsque nous expliquerons le mécanisme de la

contraction.
Notons enfin que des catégories de fibres intermédiaires ont été retenues et
appelées F II a,b et c. A n'en pas douter, ces sous classes sont l'expression d'un
phénomène continu sous-jacent. Nous verrons, en effet, qu'au niveau biochimique
et physiologique, un continuum naturel existe certainement entre les deux types
de fibres I et II (Greenhaff P, Timmons J, 1998). C'est par simple commodité
que nous maintenons la classification que nous venons de rapporter.
Voyons à présent comment les fibres se répartissent dans le corps humain.
2.2 Répartition des types de fibres
Les proportions des différentes fibres varient d'une personne à l'autre pour un
même groupe musculaire et d'un groupe musculaire à l'autre chez la même
personne.
Répartition en fonction des muscles
Les pourcentages de fibres lentes et rapides ne sont pas les mêmes dans tous les
muscles du corps. Par exemple, le muscle soléaire situé dans le mollet sous les
muscles jumeaux est presque complètement composé de fibres lentes chez tous
les sujets. En revanche, le quadriceps situé sur l'avant de la cuisse est souvent
de composition mêlée.
En dépit de l'hétérogénéité de la répartition des fibres dans les muscles du
corps, les muscles des bras et des jambes d'une même personne ont en général
des compositions semblables en fibres. Avec l'âge, nos muscles tendent à perdre
des fibres rapides au profit des fibres lentes.
Répartition en fonction des personnes
Selon les personnes, la proportion de fibres rapides peut varier entre 20 et
80%. Ajoutée aux données génétiques, la pratique permet de rendre compte de
cette répartition.
2.3 Type de fibres et force développée
Selon Denis et Lacour (1979), l'aptitude d'un individu à fournir un travail
supramaximal est en partie déterminée par la proportion de fibres à contraction
rapide ou lente dans ses différents groupes musculaires. La force maximale que
peut développer un groupe musculaire est directement corrélée à sa teneur en
fibres de type II. Il n'est pas étonnant de constater que la spécialisation dans
une discipline est associée à une proportion élevée de fibres spécifiques de la
qualité sollicitée. En moyenne, les coureurs de 100m possèdent 75% de fibres II
alors que les marathoniens n'en disposent que de 20%.
Ce constat s'applique aux sujets entraînés. Il se limite à une corrélation et ne
saurait être assimilé à un déterminisme simple. Pour être plus clair, disons que ce
n'est pas parce que nous disposons de beaucoup de fibres rapides que nous
serons forcément bons sur 100m. Dans les exercices explosifs, les performances
ne sont pas corrélées au pourcentage de fibres rapides. Ceux qui sont bons ont
des fibres rapides mais ceux qui ont des fibres rapides ne sont pas forcément

bons. En fait, les performances sont fortement influencées par les qualités
techniques, de coordination (contraction - relâchement), de force maximale.
Nous venons de dire qu'un muscle composé de nombreuses fibres rapides est
capable de produire plus de force et plus vite qu'un muscle qui en serait
dépourvu. Partant de ce constat, nous pensons être en droit de passer de la
pluralité à l'unité. Nous pensons être en droit de penser qu'une seule fibre
rapide est capable de produire plus de force qu'une fibre lente. En est-il bien
ainsi ?
Nous savons que les fibres lentes (I) stimulées développent leur tension
maximale en 110ms contre 50ms pour les fibres rapides (II). Une fibre II est
donc plus rapide qu'une fibre I. En revanche, en 1967, R. Close a montré que
fibres lentes (I) et rapides (II) prises individuellement à leur niveau de tension
maximale, ne diffèrent guère quant à la force qu'elles développent.
Si cette observation est vraie, si la force pouvant être produite par une fibre
rapide ne diffère pas sensiblement de celle dispensée par une fibre lente, qu'est
ce qui permet d'expliquer qu'un muscle composé de fibres rapides produit plus
de force qu'un muscle composé de fibres lentes ? Pour le comprendre, nous
devons poser notre regard sur les fibres nerveuses.
3 Le regard sur les nerfs
Nous voilà, à présent, à l'interface entre l'innervation, c'est à dire la commande
du muscle par le nerf et la force appliquée sur l'environnement. Débutons notre
étude par une description de l'innervation du muscle.
3.1 Organisation de l'innervation
Qu'observons-nous lorsque nous raisonnons au niveau du rapport entre les nerfs
et les fibres musculaires ?
Un seul nerf innerve plusieurs fibres. Le nerf est un câble qui, au niveau de sa
partie finale, se divise en de nombreuses terminaisons. Chaque prolongement se
termine au niveau d'une fibre musculaire. L'ensemble composé de la fibre, de ses
prolongements et des fibres musculaires associées prend le nom d'unité
motrice.
Une unité motrice est donc constituée d'un seul neurone et des différentes
fibres musculaires qu'il innerve. Lorsqu'on abordera le recrutement des fibres
par les voies nerveuses, c'est au niveau des unités motrices que nous devrons
raisonner. C'est elle qui va nous permettre de comprendre pourquoi les unités
motrices rapides sont plus fortes que les lentes.
3.2 Innervation et force développée
Les fibres lentes sont innervées par un motoneurone (neurone moteur) à petit
corps cellulaire. Ce motoneurone innerve un groupe de 10 à 180 fibres. Une unité
motrice de type rapide a un corps cellulaire important et innerve de 300 à 800

fibres musculaires. Un gros motoneurone provoque donc la contraction de plus de
fibres qu'un petit motoneurone. Cette différence d'innervation est
certainement à l'origine des écarts de force développée par les deux sortes de
fibres.
Les unités motrices rapides produisent plus de force que les unités motrices
lentes parce que chaque unité motrice rapide contient plus de fibres musculaires
qu'une unité motrice lente ! Rappelons que chaque fibre prise individuellement
produit approximativement la même force. Cette donc la somme des fibres
activées qui explique le gain de force. Répétons le ; la force développée dépend
du nombre de fibres recrutées et non pas du type de fibre.
Voyons plus en détail les mécanismes de ce recrutement.
Le recrutement des fibres
Le recrutement des fibres se fait selon la loi du tout ou rien. Rien ne se passe
tant que les influx nerveux n'ont pas atteint un seuil limite. Dès que ce seuil
liminal est dépassé, une stimulation maximale est observée au niveau de la fibre
musculaire. Comme toutes les fibres musculaires d'une même unité motrice
reçoivent la même stimulation nerveuse toutes se contractent au maximum.
Lorsqu'une force faible est nécessaire pour le mouvement, seules quelques
fibres sont excitées. L'augmentation de force se fait par augmentation du
nombre de fibres recrutées au maximum de leur force.
Dans les années 1970, Gollnick et Coll ont montré que le recrutement sélectif est
déterminé non par la vitesse de contraction mais par le niveau de la force à
développer (Gollnick et Hodgson, 1986 ; Gollnick, Piehl, Saltin 1974). Les fibres
rapides IIb sont difficiles à solliciter. Le recrutement est progressif, des lentes
aux rapides. C'est ce que pose la théorie du recrutement ordonné des fibres
musculaires ; théorie toutefois peu robuste. Pour une force donnée ce seraient
toujours les mêmes unités motrices qui seraient recrutées, les autres attendant
leur tour, à savoir une augmentation de la force à appliquer sur l'environnement.
Contrairement à ce nous pourrions penser, lors d'efforts maximaux, le système
nerveux ne recrute pas 100% des fibres disponibles. Seules 50 à 70% des unités
motrices seraient activées. Ce constat signifierait que 30 à 50% des fibres ne se
contractent jamais. Nous pouvons nous demander pourquoi l'organisme prend de
telles précautions. Une hypothèse de travail pourrait être que des fibres
seraient mises en réserves. Elles prendraient le relais au cas où les fibres
actives seraient épuisées. Par ailleurs, cette prudence permettrait d'éviter des
lésions musculaires ou tendineuses. Si au même moment nous pouvions contracter
toutes les fibres d'un muscle, la force ainsi générée pourrait probablement
déchirer le muscle et ses tendons.
Le modèle qui ressort du fonctionnement que nous venons de décrire est donc le
suivant. Dans toutes les activités, le système nerveux recrute
préférentiellement les fibres les mieux adaptées à la production du niveau de
puissance exigé. Au fur et à mesure de l'exercice, les fibres se fatiguent
(manque de glycogène, accumulation de déchets...) et le système nerveux doit
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%