3 - AMPCfusion

Parcours 3 Physiologie de la
reproduction
cours n°3
21/10/2015
J. Cohen-Tannoudji
RT : DESCOURBES Benjamin
RL : COMBIER Marie
L’axe hypothalamo-hypophyso-gonadique
Plan :
I. Les fonctions des gonades
A. Production de gamètes
1) Production chez le mâle
2) Production chez la femelle
B. Synthèse d’hormones stéroïdes
1) Nature des stéroïdes produits
2) Évolution au cours de la vie et cibles d’action
II. Le contrôle neuroendocrine et endocrine des fonctions gonadiques
A. Organisation fonctionnelle du complexe hypothalamo-hypophysaire
B. La GnRH hypothalamique
1) Neurones hypothalamiques
2) Régulation de la sécrétion
C. Les hormones gonadotropes hypophysaires
III. Mutations de gènes clés et pathologies de la fonction de reproduction
A. Mutations activatrices: exemple du récepteur de la LH
B. Mutations inactivatrices: exemple de la sous-unité β de la FSH
Introduction
Nous nous intéressons dans ce cours à l’axe hypothalamo-hypophyso-gonadique. Cet axe régule
la reproduction. Le contrôle central se fait grâce à la GnRH, qui va agir sur les cellules
gonadotropes de l’adénohypophyse qui vont produire
LH et FSH agissant sur les gonades pour réguler la
gamétogénèse ainsi que la stéroïdogenèse. Il y a
ensuite un rétrocontrôle de cette production.
Mais quand il y a des perturbations de cet axe, quelles
sont les conséquences sur la reproduction ?

I. Les fonctions des gonades
A. Production de gamètes
Les gonades ont 2 principales fonctions. La 1ère est la production de gamètes (fonction
exocrine), la 2ème est la production d’hormones stéroïdiennes et de facteurs non stéroïdiens
(fonction endocrine) qui ont une action locale pour réguler la gamétogénèse mais aussi une
action plus large sur l’organisme (comme voies génitales, SNC, différenciation sexuelle primaire
ou secondaire…)
1) Production chez le mâle
L’organisation de la gonade mâle est celle d’un testicule tubulaire (il n’est pas tubulaire chez les
reptiles par exemple). On le dit tubulaire car chez les mammifères il est constitué à 80% de
tubes séminifères. Le gamète masculin est le spermatozoïde. La spermatogenèse se fait dans les
tubes séminifères. On trouve aussi dans ces tubes les cellules de Sertoli. La production de
spermatozoïdes est massive et continue (elle ne s’arrêtera jamais) à partir de la puberté. La
production endocrine a, elle, lieu dans le tissu interstitiel où l’on trouve les cellules de Leydig.
2) Production chez la femelle
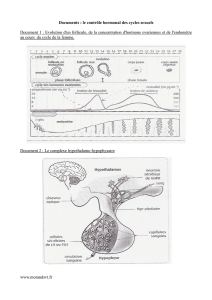
Chez la femelle c’est différent, le gamète est l’ovocyte, qui se trouve au sein du follicule ovarien.
Il est difficile de dissocier folliculogenèse d’ovogenèse. L’entrée en méiose est beaucoup plus
précoce chez la femelle que chez le mâle puisque chez celle-ci elle commence dès la vie
embryonnaire. À la naissance (ou peu de temps après), le stock définitif de cellules germinales
est fait. Les follicules primordiaux sont en périphérie. Dans ces follicules, l’ovocyte est bloqué au
stade de prophase I (on parle ici d’ovocyte I), en effet la méiose est discontinue.
Pour 99% des follicules ovariens, il y a un phénomène de dégénérescence, c’est l’atrésie
folliculaire. A la ménopause, il n’y a plus de dialogue harmonieux entre les ovaires et
l’hypothalamus et l’hypophyse. Elle peut arriver tôt on parle alors d’IOP (Insuffisance Ovarienne
Prématurée).
Les follicules qui continuent d’évoluer sont de plus en plus gros jusqu’au stade de follicule pré-
ovulatoire. La croissance se fait majoritairement grâce aux cellules de la granulosa. L’ovocyte qui
sera ovulé débloquera sa méiose pour se re bloquer en métaphase II.
Il y a un fonctionnement cyclique de la puberté à la ménopause. Le cycle ovarien est composé
d’abord de la phase folliculaire (14-15jours) suivie de l’ovulation et enfin de la phase lutéale (14-
15j). La phase folliculaire est la phase de croissance du follicule vers le follicule pré-ovulatoire et
a pour stéroïde majeur l’estradiol. La phase lutéale permet la mise en place du corps jaune et a
pour stéroïde principal la progestérone (chez la femme il y a quand même de l’estradiol mais de
façon moindre). Le cycle sexuel n’a pas lieu que sur les ovaires mais aussi sur les voies génitales.
On parle de cycle menstruel chez les Hommes et les primates de l’ancien monde et on parle de
cycle oestrien chez la majorité des mammifères.

B. Synthèse d’hormones stéroïdes
1) Nature des stéroïdes produits
Il existe 3 grandes classes de stéroïdes grâce aux cytochromes P450 (CYP450) et StAR: les
progestagènes, les androgènes et les oestrogènes. Les stéroïdes dérivent du cholestérol. La
première étape est l’entrée du cholestérol dans la mitochondrie grâce à la protéine StAR (entre à
la membrane interne), c’est l’étape limitante. Dans la mitochondrie, le cholestérol est métabolisé
en prégnénolone qui sort ensuite dans le réticule lisse pour continuer sa maturation. A la fin de
la maturation, les androgènes peuvent être pris en charge par l’aromatase pour être transformés
en oestrogènes. Chez l’homme, une partie du rétrocontrôle a donc besoin des récepteurs aux
oestrogènes hypothalamiques. Ces modifications périphériques des androgènes peuvent
modifier leur action. Chez la femme, il y a des concentrations différentes en fonction du moment
du cycle.
2) Evolution au cours de la vie et cibles d’action.
Chez le mâle, il y a un pic de production de testostérone durant la vie foetale induisant une
masculinisation des organes génitaux externes (aussi rôle de l’AMH). La concentration de
testostérone sera ensuite très basse. En période pré-pubertaire on voit réapparaître les cellules
de Leydig (grâce à l’activation des neurones à GnRH) ce qui va permettre l’augmentation de la
concentration de testostérone et induire le développement des organes génitaux externes, des
voies sexuelles, une virilisation, l’établissement de la fertilité, etc. La testostérone permet
ensuite le maintien de la fonction sexuelle durant la vie sexuelle (et oui, logique).II. Le contrôle
neuroendocrine et endocrine des fonctions gonadiques.
II. Le contrôle neuroendocrine et endocrine des fonctions
gonadiques
A. Organisation fonctionnelle du complexe hypothalamo-hypophysaire

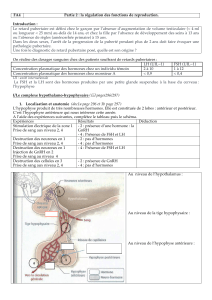
L’hypothalamus se trouve dans le diencéphale (au niveau du plancher et du bord du 3eme ventricule).
Il est prolongé par l’hypophyse et donne la tige pituitaire, l’éminence médiane et aussi la
neurohypophyse. L’hypophyse est faite de 2 parties. La 1ère est située derrière c’est la post-hypophyse
ou neurohypophyse (dérive du neurectoderme) car elle est exclusivement nerveuse (elle contient les
axones des neurones hypothalamiques à ocytocine et vasopressine, ainsi que des cellules gliales). La
2ème est située devant, c’est l’adénohypophyse ou antéhypophyse et c’est elle qui va nous intéresser
dans ce cours. C’est une glande endocrine avec un grand mélange de cellules différentes
(gonadotropes, thyréotropes, etc).Il n’y a que 10 à 15% de cellules gonadotropes dans
l’adénohypophyse.
Il n’y a pas de lien direct entre l’hypothalamus et l’adénohypophyse, c’est pourquoi il faut un sytème
pour libérer la GnRH de l’hypothalamus à l’hypophyse: c’est le système porte. Le premier plexus se
trouve dans l’éminence médiane et c’est là que sera libérée la GnRH. La GnRH va ensuite sortir au
niveau du 2ème plexus, qui se situe dans l’adénohypophyse, pour agir sur les cellules gonadotropes
qui pourront libérer LH et FSH.
B. La GnRH hypothalamique
1) Neurones hypothalamiques
Les neurones à GnRH sont les seuls produits hors du SNC et sont trouvés très précocément dans une
structure embryonnaire olfactive, la placode olfactive. Ils doivent ensuite migrer jusqu’à
l’hypothalamus. Une fois « arrivés », il y a une différenciation terminale qui vise a emmener leur
axone à l’éminence médiane afin que la GnRH puisse être libérée dans le système porte. Il y a 800 à
1000 neurones à GnRH chez l’Homme, qui ont une localisation diffuse (surtout hypothalamus médian
et antérieur, mais aussi dans structures extra-hypothalamiques et extra-cérébrales).
Quand il y a un défaut de migration on trouve un hypogonadisme hypogonadotrope souvent associé à
une anosmie, c’est le syndrome de Morsier-Kallmann.
La séquence de la GnRH de type I (retrouvé chez TOUS les mammifères) est un décapeptide:
pGlu-His-Trp-Ser-Tyr-Gly- Leu-Arg-Pro-Gly-NH2.
La sécrétion de GnRH est pulsatile, et est toujours suivie d’une sécrétion de LH. Ces pulses
permettent d’éviter une désensibilisation. Les changements de rythme de ces pulses
déterminent les grandes étapes de la puberté.
2) Régulation de la sécrétion
Les neurones à GnRH sont le centre de contrôle d’un grand nombre d’informations. Ils ont une
action différente en fonction des signaux stéroïdiens et des autres signaux internes ou
environnementaux. Cependant ces neurones à GnRH ont très peu de récepteurs. L’idée est venue
que l’action se fait plutôt en amont sur des neurones afférents.

On a découvert le système kisspeptines/GPR54. GPR54 est un récepteur qui lie les peptides
dérivés de la messine (gène Kiss1). Ce récepteur se trouve dans le cerveau, le placenta et
l’hypophyse. Lorsque le gène à l’origine de GPR54 est muté il n’y a pas de puberté ainsi qu’un
hypogonadisme hypogonadotrope. De plus, une administration de kisspeptines entraine une
sécrétion de GnRH. Il y a expression de GPR54 dans les neurones à GnRH. Kiss a un rôle central
dans la régulation de l’activité des neurones à GnRH.
Chez l’homme et la femme il y a un rétrocontrôle négatif grâce aux stéroïdes (récepteurs aux
androgènes et aux oestrogènes après aromatisation) principalement au niveau de
l’hypothalamus sur les neurones à GnRH.
Chez la femme lors de l’ovulation il y a un rétrocontrôle positif dû à une très grande
augmentation de l’estradiol durant un temps prolongé. Normalement les neurones Kiss sont
inhibés par l’estradiol mais chez la femme certains sont activés grâce à l’estradiol.
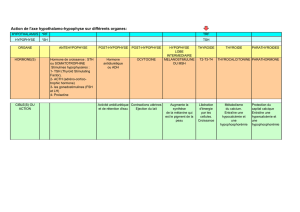
3) Les hormones gonadotropes
Les hormones gonadotropes sont des hormones glycoprotéines à 2 sous-unités. Il existe 4types
d’hormones glycosylées: la FSH, la LH, la TSH et la CG. Elles ont toutes la même chaine α mais pas
la même chaine β qui fait leur spécificité (ce sont donc des hétérodimères).
Les récepteurs à LH et FSH sont des récepteurs couplés à une protéine G (RCPG) couplés à la
voie de l’adénylate cyclase/AMPc. Le récepteur à la LH se trouve sur les cellules de Leydig et
induit la stéroïdogenèse et le récepteur à FSH sur les cellules de Sertoli et induit la
gamètogenèse.
Chez la femme, l’initiation et le début de maturation du follicule n’ont pas besoin de FSH et de LH
(jusqu’au petit follicule à antrum). Cependant FSH et LH sont indispensables à la maturation
terminale.
III. Mutations de gènes clés et pathologies de la fonction de
reprouction
On trouvera ici les mutations activatrices (gain de fonction), dominantes à l’état hétérozygote, et
les mutations inactivatrices (perte de fonction), à transmission autosomique récessive.
A. Mutations activatrices: exemple du récepteur de la LH
 6
7
6
7
1
/
7
100%