Goupil_Mathieu_2014_these - Papyrus : Université de Montréal

Université de Montréal
Analyse des altérations de l'immunité T-dépendante à
l'égard de Candida albicans chez la souris transgénique
exprimant le génome du VIH-1
par
Mathieu Goupil
Département de microbiologie, infectiologie et immunologie
Faculté de Médecine
Thèse présentée à la Faculté de médecine
en vue de l’obtention du grade de PhD
en microbiologie et immunologie
Novembre 2013
© Mathieu Goupil, 2013

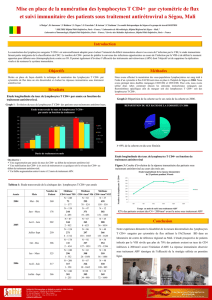
Résumé
La candidose oro-pharyngée (COP) est l’infection fongique opportuniste la plus
commune chez les individus infectés par le VIH-1. La production des cytokines Il-17 et Il-22
par les lymphocytes Th17 est importante lors de la résolution de la COP, puisque ces
cytokines induisent la production de peptides antifongiques et le recrutement des neutrophiles
polymorphonucléaires. Toutefois, les lymphocytes Th17 sont préférentiellement déplétés chez
les individus infectés par le VIH-1. Le modèle de COP chez la souris transgénique (Tg)
CD4C/HIVMutA, exprimant les gènes nef, env et rev du VIH-1, permettra de déterminer si des
altérations quantitatives et/ou fonctionnelles des sous-populations de lymphocytes T CD4+
causent la sensibilité à la candidose. Les sous-populations Th1, Th2, Th1Th17, Th17 et Treg,
ainsi que leurs précurseurs, les lymphocytes T CD4+ naïfs, sont sévèrement déplétées dans les
ganglions cervicaux de la souris Tg. Cependant, les lymphocytes T CD4+ naïfs conservent la
capacité à se différencier in vitro en présence de cytokines polarisantes et à produire les
cytokines typiques des diverses sous-populations. De plus, les cytokines requises pour la
polarisation des lymphocytes T CD4+ naïfs n’étaient pas réduites dans les ganglions cervicaux
des souris Tg, 7 jours après le début de l’infection. Les gènes S100a8, Ccl20, Il17 et Il22
étaient surexprimés en réponse à la COP chez la souris non-Tg, mais pas chez la souris Tg. Le
traitement de souris Tg infectées à l’aide de la combinaison des cytokines Il-17 et Il-22 réduit
significativement la charge fongique buccale de C. albicans et le nombre d’hyphes dans
l’épithélium de la langue et restaure la capacité à surexprimer des gènes S100a8, Ccl20 et Il22.
Ces résultats démontrent que la perturbation de l’induction de l’immunité innée par l’Il-17 et
l’Il-22 augmente la susceptibilité à la COP chez la souris Tg.
Mots-clés: Candidose oro-pharyngée, cytokine, Il-17, Il-22, modèle animal, Th17, lymphocyte
T régulateur, VIH-1.

Abstract
Oropharyngeal candidiasis (OPC) is the most common opportunistic fungal infection in
HIV-infected individuals. OPC resolution involves Il-17 and Il-22 production by Th17 cells
through oral antifungal peptide production and polymorphonuclear neutrophil recruitment.
Conversely, Th17 cells are preferentially depleted in HIV-infected individuals. The OPC
model in transgenic (Tg) mice expressing nef, env and rev from the HIV-1 genome enables the
study of the quantitative and functional defects of the CD4+ T-cell subpopulations. The Th1,
Th2, Th1Th17, Th17 and Treg subpopulations, as well as naïve CD4+ T-cell precursors, are
severely depleted in the cervical lymph nodes (CLNs) of Tg mice. However, the
differentiation capacity of naïve CD4+ T-cells in response to polarizing cytokines was
maintained in vitro in Tg mice, as well as their ability to produce the signature cytokines of the
various subpopulations. Moreover, the polarizing cytokines were not reduced in the CLNs of
Tg mice, 7 days after infection. The S100a8, Ccl20, Il17 and Il22 genes were up-regulated in
response to OPC in non-Tg mice, but not in Tg mice. Treatment of infected Tg mice with a
combination of Il-17 and Il-22 cytokines significantly reduced the oral fungal burdens of C.
albicans as well as the number of hyphae in the tongue epithelium. Treatment also restored
S100a8, Ccl20 and Il22 up-regulation in Tg mice. These results show that defective induction
of innate immunity, normally mediated by Il-17 and Il-22, increases the susceptibility to OPC
in these Tg mice.
Keywords: Animal model, cytokine, HIV-1, Il-17, Il-22, oropharyngeal candidiasis,
regulatory T-cell, Th17.

Table des matières
Liste des tableaux ..................................................................................................................... vi
Liste des figures ...................................................................................................................... vii
Liste des sigles et abréviations ................................................................................................ ix
Remerciements ....................................................................................................................... xii
Chapitre 1 - Candida albicans et la candidose ........................................................................ 1
Candida albicans ........................................................................................................................ 1
Épidémiologie ............................................................................................................................. 5
Manifestations cliniques ............................................................................................................. 6
Traitement ................................................................................................................................. 10
Pathogénèse ............................................................................................................................... 11
Chapitre 2 - Immunité contre Candida albicans .................................................................. 17
Les protéines salivaires ............................................................................................................. 17
La muqueuse buccale ................................................................................................................ 22
Kératinocytes ............................................................................................................................ 22
Neutrophiles polymorphonucléaires (PMNs) ........................................................................... 23
Cellules NK (Natural Killer) .................................................................................................... 24
Cellules de Langerhans ............................................................................................................. 25
Macrophages ............................................................................................................................. 26
Lymphocytes B ......................................................................................................................... 28
Lymphocytes T ......................................................................................................................... 28

iv
Chapitre 3 – Le virus de l’immunodéficience humaine ....................................................... 35
Épidémiologie du VIH .............................................................................................................. 35
VIH-1 ........................................................................................................................................ 36
Infection au VIH- ...................................................................................................................... 38
Traitement ................................................................................................................................. 43
Chapitre 4 – Modèles d’études de la candidose oro-pharyngée .......................................... 45
Hypothèse ................................................................................................................................. 51
Chapitre 5 – Matériel et méthodes ........................................................................................ 53
Souris transgénique CD4C/HIVMutA ......................................................................................... 53
Modèle de candidose oro-pharyngée chez la souris transgénique CD4C/HIVMutA .................. 53
Analyse des sous-populations de lymphocytes T CD4+ par la cytométrie en flux ................... 54
Tri cellulaire de lymphocytes T CD4+ naïfs ............................................................................. 56
Différenciation cellulaire de lymphocytes T CD4+ naïfs in vitro ............................................. 57
Production de cytokines in vivo dans les ganglions cervicaux ................................................. 58
Injection d’Il-17 et/ou d’Il-22 ................................................................................................... 59
Analyses statistiques ................................................................................................................. 60
Chapitre 6 – Résultats ............................................................................................................ 61
Les sous-populations de lymphocytes T CD4+ sont déplétées chez la souris transgénique
CD4C/HIVMutA .......................................................................................................................... 61
L’expression des gènes polarisant la différenciation des lymphocytes T CD4+ est altérée chez
la souris transgénique CD4C/HIVMutA ...................................................................................... 63
L’expression du transgène VIH-1 n’altère pas la production de cytokines chez les lymphocytes
T CD4+ différenciés in vitro ..................................................................................................... 65
Altération de la production de cytokines dans les ganglions cervicaux chez la souris
transgénique CD4C/HIVMutA .................................................................................................... 66
Augmentation de la résistance à la candidose oro-pharyngée suite au traitement de souris
transgéniques CD4C/HIVMutA infectées au C. albicans avec les cytokines Il-17 et Il-22 ........ 68
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
1
/
195
100%