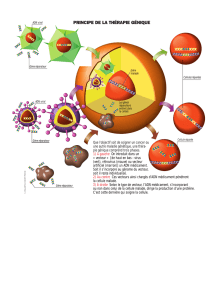
Le système du TAP tag, principe, vecteurs et souches utilisées Le

Encart 1 : Le système du TAP tag, principe, vecteurs et souches utilisées
Le système du TAP tag est une méthode de purification par affinité. Cette méthode consiste à étiqueter une
protéine dont on sait qu’elle interagit avec le complexe d’intérêt, à exprimer cette protéine dans le parasite puis,
après extraction des protéines en conditions non dénaturantes, à purifier et à analyser les protéines qui lui sont
associées. Nous avons choisi d'utiliser pour ce faire une étiquette particulière appelée TAP (Tandem Affinity
Purification) (Rigaut et al., 1999; Puig et al., 2001). Le TAP tag est maintenant couramment utilisé chez
Trypanosoma brucei et ce système a permis de purifier de nombreux complexes (pour plus de détails, voir
matériel et méthodes). Différents vecteurs ont été utilisés pour permettre le marquage des protéines d'intérêt :
(i) Le vecteur pTSARib TAP (figure 3.1A) : couramment utilisé au laboratoire (Walgraffe et al., 2005), il
permet de fusionner l'étiquette TAP au domaine carboxyterminal (C-term) de la protéine d'intérêt. Ce vecteur
contient une séquence codant pour le TAP et un gène de résistance à un antibiotique, permettant de sélectionner
les parasites qui l’ont intégré suite à la transfection. Grâce à des régions d'homologie présentes dans le vecteur,
la construction, après transfection, est intégrée dans le locus ribosomique du parasite et la protéine de fusion
résultante est surexprimée. Ce vecteur est transfecté dans des parasites sauvages.
(ii) Les vecteurs pLew82 NLS NTAP (figure 3.1B) et pLew100 NLS NTAP (figure 3.1C) permettent
l'expression inductible d'une sous-unité et son étiquetage du côté de son domaine aminoterminal (N-term). Ces
deux vecteurs sont des vecteurs d’expression inductible, contenant un promoteur de la polymérase T7 et une
séquence de l'opérateur de l'opéron Tet bactérien sensible à la doxycycline. Ces vecteurs contiennent en outre
des régions homologues aux séquences ribosomiques du parasite permettant l’insertion de la construction
d’intérêt dans ces régions du génome (Wirtz et al. 1999). La séquence du TAP tag et un signal de localisation
nucléaire (NLS= Nuclear Localisation Signal, Marchetti et al. 2000) ont été insérés dans les vecteurs pLew
pour créer les vecteurs pLew NLS NTAP. Dans le vecteur pLew 82, le gène de résistance aux antibiotiques est
sous le même promoteur que le gène d’intérêt. Les parasites sélectionnés exprimeront donc obligatoirement la
protéine d’intérêt, mais à un faible taux. La doxycycline permet d’augmenter le taux d’expression. Le vecteur
pLew100 quant à lui contient deux promoteurs différents, un pour le gène d’intérêt et un pour le gène de
résistance. Un bruit de fond d’induction n’est donc pas obligatoire pour la sélection des parasites. En outre, le
promoteur T7 responsable de l’expression du gène d’intérêt est muté, ce qui diminue le taux d’expression. Ces
vecteurs sont transfectés dans la souche 29:13 (stade procyclique) (Wirtz et al. 1999) qui contient la polymérase
T7 et le répresseur tétracycline.

Vecteur d'ARNi
Figure 3.2 : Représentation des vecteurs utilisés au cours de ce travail et permettant
l'invalidation par ARNi d'un gène d'intérêt
Tet op
Prom
Tet R
Gène
Tet R
Tet op
Prom
Prom Promoteur T7
Tet op
Opérateur de l'opéron
tétracycline
Tet R
Répresseur tétracycline
Gène Gène ou morceau
du gène d'intérêt
Encart 2 : L’ARNi, principes, vecteurs et souches utilisées
L'ARNi consiste à produire un petit ARN double brin correspondant à une portion du gène d'intérêt. Cet ARN
double brin active une machinerie enzymatique qui le clive en petits fragments de 21-23 nucléotides appelés
siRNA (Small Interfering RNA). Ces siRNA induisent la dégradation de l'ARNm correspondant, empêchant
ainsi l'expression du gène ciblé. Différents plasmides permettant d'induire ce "knock-down" spécifique ont
donc été créés.
Nous avons utilisé dans ce travail le vecteur pZJM (Wang et al. 2000) et le vecteur p2T7 tiTa 177
(Wickstead et al. 2002). Chacun d'entre eux contient deux promoteurs spécifiques de l'ARN polymérase T7,
disposés tête-bêche et entre lesquels peut être inséré le fragment du gène d'intérêt. Ces promoteurs sont eux-
mêmes sous le contrôle de l'opérateur de l'opéron tétracycline bactérien sensible à la doxycycline (figure 3.2).
Les vecteurs contenant le fragment du gène à invalider sont transfectés dans des souches de trypanosomes
spécifiques qui expriment l'ARN polymérase T7 ainsi que le répresseur Tet. Lorsqu'on ajoute la doxycycline au
milieu de culture contenant les parasites, la transcription de l'ARN double brin (ARNdb) s'effectue, le système
d'ARNi se met en place et les transcrits du gène d'intérêt sont dégradés.
Les souches de parasites utilisées sont la souche 29.13 (Wirtz et al. 1999) au stade procyclique et la souches
328.114 (Wirtz et al. 1999) au stade sanguicole. Ces deux souches de parasite expriment la polymérase T7 ainsi
que le répresseur tétracycline. Le vecteur pZJM contient des régions d'homologie avec l'ADNr permettant son
insertion au sein du locus ribosomique tandis que le vecteur p2T7-177 est inséré dans les minichromosomes.
Ceux-ci présentent une cible clef pour des vecteurs inductibles car ils ne sont jamais transcrits (voir
introduction). L'utilisation du vecteur p2T7 permet donc de diminuer le bruit de fond d'induction, fréquemment
observé en absence de doxycycline avec le système d'invalidation pZJM, et qui peut être problématique lorsque
le gène considéré est indispensable pour la survie du parasite. Le vecteur p2T7 n'a cependant été utilisé dans ce
travail que sur des trypanosomes au stade sanguicole car, en raison des résistances spécifiques des parasites aux
drogues de sélection, seule la souche 328.114 (sanguicole) était compatible avec le vecteur dont nous
disposions.

Encart 3: Analyse de l’effet de l’ARNi sur la transcription
L'analyse de la quantité d'ARN au sein des cellules invalidées par ARNi a été effectuée par
Northern blot.
L'analyse des transcrits primaires a été effectuée en itilisant trois types de techniques
(i) Les "run-on" in situ visualisés par immunofluorescence. Cette technique développée par
l'équipe de Gull (Navarro et al. 2001) permet de visualiser la transcription en cours dans la
cellule. Brièvement, des parasites vivants, invalidés ou non pour le gène d'intérêt, sont
perméabilisés et incubés dans un milieu permettant la transcription. Ce milieu contient les
différents nucléotides dont certains sont marqués avec le bromouridine triphosphate (BrUTP). Les
ARN nouvellement synthétisés contiennent donc le nucléotide modifié et peuvent être visualisés
par immunofluorescence à l'aide d'un anticorps dirigé contre cette molécule. Dans la suite du
manuscrit, cette technique sera appelée "run-on" in vivo.
(ii) Les marquages métaboliques visualisés par "dot-blot" (Ullu et al. 1990). Contrairement à la
technique précédente qui donne une vision globale de la transcription par immunofluorescence, le
"run-on sur dot-blot" permet de s'intéresser à la transcription primaire d'un gène particulier. Dans
ce cas-ci, l'ARN est synthétisé en présence d’un nucléotide radioactif puis utilisé comme sonde
pour s’hybrider à une séquence d'ADN, préalablement transférée sur un filtre de nitrocellulose par
"dot-blot".
(iii) Le "run-on" (Murphy et al. 1987). Cette dernière technique consiste à isoler les noyaux avant
de marquer avec de la radioactivité les ARN en cours de transcription. Ces ARN seront alors
utilisés comme sondes pour hybrider des séquences d'ADN, préalablement transférées par
Soutern blot sur filtres de nitrocellulose.
Les différentes techniques utilisées et la comparaison des résultats obtenus permettront d'avoir
une idée globale des modifications de l'activité transcriptionnelle au cours de l'invalidation de
chaque gène considéré.
1
/
3
100%