V - La fonction digestive

Par Krys3000 (Groupe « The Trust » - http://www.cours-en-ligne.tk/) Page 1
GRANDES FONCTIONS ANIMALES
CHAPITRE IV : LA FONCTION DIGESTIVE
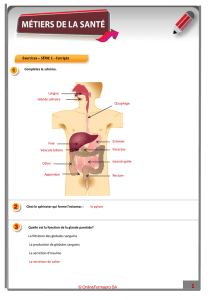
I – LA SÉCRÉTION SALIVAIRE
A lieu dans la bouche grâce à des sucs. Cette sécrétion est contrôlée par 3 paires de glandes : les glandes parotides, les glandes
sous-maxillaires, et les glandes sublinguales. Ces glandes sont constituées de lobes, qui eux-mêmes comprennent des lobules,
amas cellulaires composés par des acini (sing : acinus), groupes de cellules sécrétrices. Ceux-ci sont reliés entre eux via des
canaux intralobulaires (entre 2 acini seulement), intercalaires (entre plusieurs acini) et extralobulaires (menant la sécrétion dans
son lieu final d’action)
Les Acini eux-mêmes existent sous trois formes, une pour chaque glande :
- Glandes sublinguales : Le type d’acinus associé est un acinus muqueux, composé de cellules muqueuses, qui
contiennent des flocons de mucines. Son canal extralobulaire est en fait une multitude de canaux qui constituent le
palais buccal. 4 % de la sécrétion basale.
- Glandes parotides : Le type d’acinus associé est un acinus séreux, composé de cellules séreuses, qui contiennent des
granules de zymogène fournissant de l’amylase (bloquée par atropine et stimulée par pilocarpine). Son canal
extralobulaire est le canal de Stenon (aboutit au niveau de la 2
ème
molaire supérieure). 25 % de la sécrétion basale.
- Glandes sous-maxillaires : Le type d’acinus associé est un acinus mixte, composé des deux types de cellules. Son canal
extralobulaire est le canal de Wharton (aboutit au niveau de la face interne du maxillaire inférieur). 71 % de la sécrétion
basale.
Il se forme 0.5 à 1.5 L de salive par jour. Elle est hypotonique mais la tonicité de la salive augmente légèrement lorsqu’il y a
sécrétion, et devient légèrement alcaline (pH maximum de 8 car régulation par ion bicarbonate). Le pH régule l’activité des
enzymes salivaires et des protéines de la salive :
• L’amylase, qui coupe les liaisons 1-4 des glucides, libérant glucose ou maltose (produit par les parotides). Fortement
dépendante du pH, elles deviennent inactive dans l’estomac (pH trop acide) mais peuvent fonctionner de nouveau dans
l’intestin.
• Mucines qui produisent du mucus pour lubrifier la bouche et aider à la déglutition et à la protection de la muqueuse
gastrique
• RNAses, DNAses, lipases
• Albumine, Immunoglobulines (protéines plasmiques)
• Facteurs de croissance comme le NGF
• Protéines inhibitrices et analgésiques

Par Krys3000 (Groupe « The Trust » - http://www.cours-en-ligne.tk/) Page 2
La sécrétion salivaire est sous contrôle du système nerveux autonome parasympathique mais également un peu du système
endocrinien. Toutefois, elle ne peut avoir lieu que si le SNA Orthosympathique n’est PAS sollicité, c'est-à-dire lorsqu’il n’y a pas
de danger. L’orthosympathique, toutefois, active la production de mucine rendant la salive plus épaisse.
Le contrôle se fait au travers des nerfs crâniens trijumeau (V), glossopharyngien (IX) et vague (X) pour l’afférence, et le facial
(VII) et glossopharyngien pour l’efférence. L’acétylcholine et la substance P servent de neuromédiateurs. Cela se fait selon deux
réflexes :
- Réflexe absolu : déclenchés par les stimuli physiologiques (présence d’aliments dans la bouche, mouvement de la
mâchoire et de la langue, mastication, distension de l’œsophage, etc.… )
- Réflexe conditionnels : exemple du réflexe de Pavlov (association son/prise de nourriture)
La salive favorise la mastication et la déglutition. Elle aide au langage et favorise la gustation (dissolution des substances et
lavage des récepteurs de goût) et protège les lésions buccale/dentaire grâce aux Immunoglobulines.
La pathologie associée à l’insuffisance de production salivaire est l’hyposialie.
II – LA SÉCRÉTION GASTRIQUE
L’estomac est composé de plusieurs parties et la sécrétion est due à un épithélium muqueux monocellulaire. Celui-ci présente
des invaginations, les cryptes, dans lequel on trouve des glandes. Les parties sont :
- Le cardia, qui représente l’entrée dans l’estomac
- Le fundus, la partie haute et grosse de l’estomac, responsable de la sécrétion acide, synthétisant de l’HCl, du mucus, de
la pepsine (clive les aromatiques de façon incomplète, pepsinogène = précurseur, transformation lorsque le pH est
acide) et des Facteurs Intrinsèques (pour l’absorption de la vitamine B12 nécessaire à la maturation des globules rouges
– le complexe FI-B12 est absorbé dans l’Iléon)
- L’antre, qui constitue la partie finale horizontale, qui synthétise du mucus et de la gastrine (régulant la sécrétion acide)
- Le pylore, sortie de l’estomac, qui donne sur le duodénum.
Les sous-couches stomacales présentes les neurones permettant la motricité de la digestion.
L’épithélium de la cavité gastrique comprend 4 types de cellules :
Muqueuses : produisent le mucus (grains de mucus) en libérant du HCO
3
-
. La barrière formée par leur mucus est
épaisse de 200 µm et permet un pH relativement neutre.
Pariétales : On les retrouve dans le fundus surtout. Elles sécrètent les H
+
et les facteurs intrinsèques. Elles ont
beaucoup de mitochondries.
Principales : On les retrouve dans le fundus, elles produisent la Pepsine avec leurs grains de zymogènes.
Endocrines : Il en existe 5 types différents
o G : produisent la gastrine (antre)
o Entérochromaffines : produisent la sérotonine
o ECL ou H : produisent l’histamine (fundus)
o A : produisent l’entéroglucagon (fundus)
o D : produisent la somatostatine, inhibitrice de la sécrétion.
Les cellules muqueuses se renouvellent tous les 2 à 3 jours, les glandes elles mettent plutôt autour de quelques mois. La
régénération est stimulée par la gastrine.
Le volume de suc gastrique varie de 1L à 1.5 L par jour.
Lors de la sécrétion, il y a augmentation de la composition en ion : le suc est toujours hyperosmotique par rapport au plasma, et
contient donc une forte concentration de H
+
, mais paradoxalement, également de HCO
3
-
. Tous les électrolytes, en règle
générale, augmentent avec le débit, sauf le sodium qui chute. On émet deux théories pour ces variations : La théorie du double
composant mettant en jeu un composant pariétal (CO
2
+ H
2
O donne H
2
CO
3
par l’action d’Anhydrase Carbonique, ce produit se
dissociant en H
+
+ HCO
3
-
) et un non-pariétal (le H
+
est utilisé par l’ATPase consommant 1 ATP en échangeant un K
+
de l’extérieur.
Il y a alors excès de HCO
3
-
faisant entrer du Cl
-
), et la théorie du simple composant mettant en jeu une sécrétion primaire
isoosmotique.
L’estomac ne digère que 20 % des protéines ingérées lors du repas, le reste étant le travail du pancréas.
La sécrétion gastrique est sous contrôle du système Nerveux et Hormonal
• Activateurs :

Par Krys3000 (Groupe « The Trust » - http://www.cours-en-ligne.tk/) Page 3
o Hormones endocrines : gastrine, stimulant les cellules pariétales du fundus et la production de pepsine.
Surtout sécrétée dans l’antre sous 3 formes : Légère (17 aa), Big (34 aa), BBG (plus de 34 aa). Sécrétion
stimulée par acides gras et acides aminés.
o Système nerveux : Nerf vague stimulant les cellules principales et pariétales, ainsi que les cellules G
(amplification de la réponse) via un récepteur à acétylcholine de type M3.
o Hormones paracrines : histamine sécrétée dans le fundus par les cellules ECL agissant sur les cellules pariétales
pour augmenter la sécrétion de gastrine. Stimulée par l’acétylcholine du Vague et la Gastrine.
1. Inhibiteurs : H
+
(inhibent la sécrétion de gastrine - feed back), Sécrétine (hormone produite par le duodénum),
Glucagon, Somastotatine, GIP, VIP, calcitonine, prostaglandines E1/E2, EGF.
La sécrétion évolue suivant un cycle de 4-5h en 3 phases :
1. Phase céphalique (psychophysiologique) : fait intervenir les mécanismes nerveux sensoriels – la stimulation au niveau
buccal provoque une sécrétion gastrique automatique.
2. Phase gastrique : démontrée par introduction de nourriture directement dans l’estomac. Stimulation en deux
composantes – une mécanique, représentée par une distension du fundus, et une chimique, due à la présence l’aliment
dans l’antre, représentée par l’activation de la production de gastrine.
3. Phase intestinale : phase inhibitrice. Production de gastrine dans le duodénum ainsi que CCK, stimulant de façon
mineure, l’action majeure étant inhibitrice (sécrétion de sécrétine).
L’estomac sert donc de réservoir, permet la dégradation des protéines en fournissant ainsi des molécules qui seront les stimuli
de la sécrétion pancréatique. On a également un rôle sur l’hématopoïèse du fait des facteurs intrinsèques.
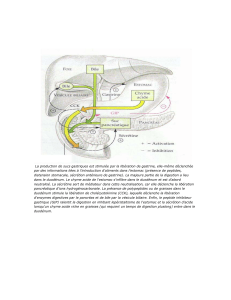
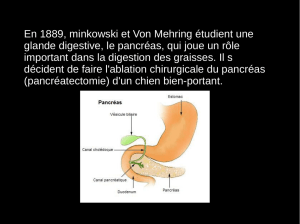
III – LA SÉCRÉTION PANCRÉATIQUE
Sécrétion exocrine. Le pancréas est composé de canaux ramifiés qui portent des acini (canaux intralobulaires entre les acini et
interlobulaires entre les lobes) drainant la sécrétion et se déversant au niveau du duodénum par des canaux collecteurs. Le
volume de suc varie de 1 à 4 L par jour.
La sécrétion pancréatique est isotonique par rapport au plasma. La concentration en HCO
3
-
est tellement haute que le pH est
basique (9). Cela est du à un échangeur H
+
/HCO
3
-
au niveau apical. Ce pH alcalin permet l’activation d’enzymes particulières
(anhydrase carbonique).
Les enzymes contenues dans le pancréas sont produites spécifiquement en fonction de la teneur des aliments.
- Amylases
- Lipases : fonctionnent avec des co-lipases. Elles coupent les triglycérides en glycérol et acides gras.
- Phospholipases : libère des lécithines
- Protéases :
o Endopeptidases coupant à l’intérieur de la protéine (autocatalyse par un système d’auto-amplification)
o Exopeptidases coupant aux extrémités des chaînes polupeptidiques
o Estérases : coupant les esters des acides ribonucléiques et désoxyribonucléiques
Le contrôle de la sécrétion pancréatique ne se fait que de façon secondaire par le SNA parasympathique, malgré le nerf X. Il est
en fait essentiellement hormonal via :
- La sécrétine, produite par les cellules S du duodénum, provoque la sécrétion pancréatique si elle est stimulée en pH
acide.
- La cholécystokinine (CCK), produite par les cellules I du duodénum et stimulée par les acides aminés et acides gras. Elle
favorise la captation des acides aminés.
- La gastrine, qui augmente la sécrétion enzymatique
- Le glucagon et la somatostatine qui servent à l’inhibition de CCK
Le SNA orthosympathique inhibe la sécrétion pancréatique.
Comme pour l’estomac, on a deux phases :
1. Phase céphalique : Elle correspond à l’infirme rôle secondaire du nerf vague (stimulation sensorielle et mécanique)
2. Phase gastrique : Fait intervenir la gastrine qui décuple la sécrétion des enzymes du pancréas
3. Phase intestinale : Déclenchée par l’arrivée des aliments dans le duodénum ; sécrétion de la sécrétine, de la CCK, d’eau
et d’HCO
3
-
ainsi que des enzymes.

Par Krys3000 (Groupe « The Trust » - http://www.cours-en-ligne.tk/) Page 4
Le pancréas sert donc à neutraliser le pH (dans l’optique de stimuler les enzymes associées) et à digérer les 80 % restant des
éléments ingérés.
IV – LA FONCTION BILIAIRE
La sécrétion de bile se fait simultanément à la sécrétion pancréatique au niveau du duodénum. Celle-ci est injectée dans une
vésicule biliaire, mais est produite par le foie, qui synthétise :
- Glucose
- Acides Aminés
- Albumine
- Immunoglobuline
Il stocke le glycogène, le fer, et les vitamines, et dégrade les protéines produisant de l’ammoniac, qui deviendra plus tard de
l’urée.
C’est un système qui va détoxifier le sang. La fonction principale est d’apporter le sang enrichi en nutriment et en toxiques et,
une fois détoxifié, le faire repartir dans la circulation par la veine hépatique.
Ce système est constitué de couches de lames de 1 ou 2 hépatocytes d’épaisseur, en étoile autour d’une veine centrale. Entre
ces lames, il y a des sinusoïdes, capillaires drainant le sang.
Le système en lui-même est drainé par les canalicules, espaces entre les hépatocytes, ainsi que par des ductules, canaux
richement vascularisés, entourés de cellules épithéliales.
Le volume de bile varie de 0.5 à 0.7 L par jour.
Cette sécrétion est isotonique par rapport au plasma. La bile est formée de sels (dérivés d’acide cholique lui-même dérivé du
cholestérol) donnant son électronégativité à la sécrétion. Elle est composée de beaucoup de HCO
3
-
. On trouve également de la
bilirubine, glucuronoconjuguée, qui permet l’absorption des aliments et la dissociation des lipides, mais cette substance doit
être éliminée dans les selles après déconjugaison en urobiline (effets neurotoxiques). Les mécanismes d’élimination se font :
- Par l’augmentation de la polarité des molécules à éliminer
- Par l’augmentation de solubilité dans l’eau
- Par l’augmentation du PM
La cholérèse (sécrétion de bile par le foie) suit le processus suivant en trois phases :
1. Sécrétion canaliculaire : les sels biliaires se concentrent dans les canalicules par osmose. Sécrétion couplée au
transport du Na
+
2. Cholérèse ductulaire et canalaire : augmentation du pH stimulée par la sécrétine
3. Hormones cholérétiques : entrée des hormones (glucagon, CCK, gastrine, histidine)
Le mécanisme d’excrétion biliaire en lui-même par la vésicule passe d’abord par une réabsorption d’eau jusqu’à 90 % du volume
initial, pour augmenter la concentration en sels, cholestérol et bilirubine. Il y a alors remplissage passif à jeun grâce à la
résistance du sphincter d’Oddi, et vidange déclenchée par la CCK (elle-même potentialisée par la sécrétine) lors du repas
(contraction de la vésicule relaxation du sphincter).
L’innervation se fait par le splanchnique (Orthosympathique) et le vague (Parasympathique).
Le rôle de la vésicule biliaire est donc de détoxifier le sang, mais aussi de solubiliser les acides gras et phospholipides grâce à sa
fonction de détergent : cela permet l’absorption par l’épithélium.
V – LA FONCTION INTESTINALE
C’est dans l’intestin que se déroule le but principal de la digestion : l’absorption d’aliments. On considère généralement le côlon
comme une partie du grêle (qui comprend duodénum, jéjunum et iléon), mais ils sont différents : si les deux possèdent bien des
repliements nommés Cryptes de Lieberkühn, seul le grêle possède des villosités sur la surface de l’épithélium. Celles-ci sont
elles-mêmes montées sur des Valvules Conniventes. Les cellules de l’épithélium intestinal sont :
- Entérocytes : Présentent des microvillosités et un glycocalyx, ainsi que des phosphatases et des Ca
2+
-ATPases
- Cellules caliciformes : produisent du mucus au rôle inconnu
- Cellules de Paneth : au fond des cryptes, sécrètent des enzymes
- Cellules souches : permettent la prolifération
- Cellules endocrines :
o Cellules S (duodénum, jéjunum) : produisent la sécrétine

Par Krys3000 (Groupe « The Trust » - http://www.cours-en-ligne.tk/) Page 5
o Cellules I (duodénum, jéjunum) : produisent la CCK
o Cellules D1 (duodénum) : produisent le GIP
o Cellules D (grêle, côlon) : produisent le VIP
o Cellules G (duodénum) : produisent de la gastrine
o Cellules EG (grêle, côlon) : produisent l’entéroglucagon
o Cellules EC (duodénum, jéjunum) : produisent la motiline
Sous l’épithélium et sur le muscle lisse se trouve un tissu conjonctif, la lamina propria. Elle supporte l’appareil vasculaire et
nerveux et contient les cellules immunitaires.
Le renouvellement cellulaire se fait dans les cryptes. Il faut 24h à peu près pour les entérocytes, le renouvellement total du tissu
prend 3 à 5 jours. Il est activé à la fois par le passage des aliments et par la flore intestinale.
Le grêle, le côlon droit, et le côlon transverse sont vascularisés par l’artère mésentérique supérieure. Il y a drainage par la veine
mésentérique supérieure se jetant dans la veine porte.
Le côlon gauche, le côlon sigmoïde, et le rectum sont vascularisés par l’artère mésentérique inférieure, et il y a drainage par la
veine mésentérique inférieure, qui donne sur la veine splénique menant à la racine gauche du tronc porte.
L’activité de digestion de l’intestin peut-être extracellulaire (pour les oligomères) par l’action d’enzymes du pancréas,
membranaire (pour les monomères) par l’action d’enzymes intestinales ou intracellulaire via l’action d’enzyme cytoplasmiques.
L’absorption est surtout intracellulaire (l’extracellulaire, qui passe par les trous dus à la desquamation, est minoritaire, et le
paracellulaire est rendu ardu par les jonctions serrées) et le transfert à l’intérieur de la membrane dépends du composé :
- Diffusion passive à travers les pores pour les petits composés
- Transport facilité (dans le sens du gradient)
- Transport actif (contre le gradient via de l’ATP)
- Transport couplé au métabolisme de la molécule transportée
- Pinocytose
Eau et sels
• Le duodénum est l’acteur d’importants transferts paracellulaires, toutefois, l’hypertonicité des nutriments envoie le
flux net du sang vers la lumière.
• Le jéjunum absorbe des glucides et des acides aminés ce qui en fait un lieu de fort potentiel osmotique : avec ses larges
pores, il absorbe 9 fois le volume d’eau absorbé par l’iléon. Il absorbera également le HCO
3
-
.
• Dans l’iléon, sera absorbé de l’eau et du sodium, et secrété du HCO
3
-
.
• Dans le côlon, 90 % de l’eau qui passe est absorbée. Le sodium est également très absorbé.
Différents facteurs peuvent agir sur la sécrétion d’eau et d’électrolytes : stimulants de la sécrétion (toxine cholérique, laxatifs),
stimulants de l’absorption (aldostérone), inhibiteurs de l’absorption.
Glucides
Dans le duodénum et le jéjunum : action d’amylases dans la lumière pour obtenir des disaccharides. Sur la surface des
entérocytes, disaccharidases (sucrase, lactase) en particulier dans le jéjunum pour transformer en monosaccharides, lesquels
sont ensuite transportés pour l’entrée de manière active (glucose, galactose) ou facilitée (fructose), et pour la sortie de manière
facilitée.
Protéines
Dans la lumière, digestion par les enzymes gastriques (pepsine) et pancréatiques (trypsine, élastase, carboxypeptidase) générant
des di-, tri-, oligopeptides pris en charge par les peptidases intestinales (aminopeptidase et dipeptidases) dans la bordure ou
dans le cytoplasme.
L’absorption passe par un transport actif coupé au sodium ou aux protons. Les Acides Aminés simples passent ensuite dans le
sang.
Lipides
Dans la lumière se trouve les lipases et les sels biliaires stimulés par la CCK des cellules I. Les lipases vont attaquer les
triglycérides, alors que les sels solubilisent les produits de l’action enzymatique et le cholestérol des aliments.
L’absorption se fait par les entérocytes : transport passif majoritaire, transport actif secondaire Na
+
dépendant pour les acides
gras à longue chaîne. A l’intérieur de l’entérocytes, on peut avoir des transformations : présynthèses dans le REL donnant
naissance à des chylomicrons après passage au Golgi, formation de VLDL en absence de graisse intestinale. Tout ça est ensuite
exocyté hors de la cellule.
Les vitamines liposolubles subissent le même type de traitement.
 6
7
8
6
7
8
1
/
8
100%