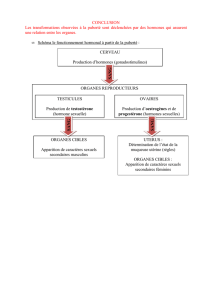
Synthèse et dynamique de lipides fusogéniques au cours de la

Synthèse et dynamique de lipides fusogéniques au cours de la libération des hormones et
des neurotransmetteurs : régulation par des protéines du retard mental
Le trafic membranaire dépend de trois mécanismes clés: le bourgeonnement de
vésicules à partir d’un compartiment donneur, le déplacement de ces vésicules vers un
compartiment accepteur, et leur fusion avec ce dernier. Au cours de l’exocytose régulée,
la fusion membranaire entraîne la libération du contenu vésiculaire dans le milieu
extracellulaire. Ce mécanisme clef permet entre autre la libération des
neurotransmetteurs et des hormones et implique des modifications de la composition
lipidiques des membranes au site d’exocytose. Nous avons démontré le rôle crucial de
certains lipides fusogéniques dans ce processus. Il s’agit maintenant de préciser leur
dynamique et voies de synthèse. Nous nous focaliserons sur l’acide phosphatidique (PA) qui
est produit spécifiquement au niveau du site d’exocytose par l’activation de la phospholipase
D (PLD). En utilisant des modèles de souris KO pour la PLD1 et la PLD2, nous
caractériserons les paramètres physiologiques altérés dans ces modèles, mais également les
effets sur les événements de libération individuels en utilisant des approches
électrophysiologiques. D’autre part, par une approche de lipidomique ciblé, nous chercherons
à caractériser les espèces précises de PA produit au niveau du site d’exocytose et
éventuellement à identifier celle(s) qui a(ont) un rôle spécifique dans la fusion membranaire.
Nous chercherons également à définir précisément la dynamique de synthèse et de conversion
du PA au cours de l’exocytose, une attention particulière à son hydrolyse en lysoPA et acide
gras dont nous détermineront l’importance possible des formes polyinsaturées.
Grace à un modèle de souris KO, nous nous concentrerons également sur la kinase RSK2 dont
la perte de fonction est responsable du syndrome de Coffin-Lowry, une pathologie associant
retard mental et défauts squelettiques. En effet, nous venons de montrer que RSK2
phosphoryle la PLD1 au cours de l’exocytose dans les cellules chromaffines. Cette étape est
nécessaire à la synthèse de PA au niveau du site d’exocytose. A partir de souris invalidées
pour RSK2, des approches électrophysiologiques seront mises en œuvre, en collaboration
avec une équipe à Bordeaux, afin de mesurer l’activité synaptique des neurones dans deux
circuits complexes, obtenus à partir de culture en tranche de cervelet et de l’amygdale. Des
mesures de la libération des catécholamines par ampérométrie, nous permettrons également
de mesurer la libération des hormones du stress à partir de cellules chromaffines murines
mises en culture. La dynamique de l’interaction entre RSK2 et la PLD1 sera étudiée par des
expériences de FRET tandis que la synthèse de PA sera étudié en temps réelle grâce à
l’emploie d’une sonde fluorescente spécifique de ce lipide. Finalement, l’importance
fonctionnelle de RSK2 dans la synthèse et la dynamique des lipides fusogéniques au cours de
la libération des hormones et des neurotransmetteurs sera comparée à celle d’autres protéines
impliquées dans des pathologies de retard mental.
Synthesis and dynamics of fusogenic lipids during neurotransmitter and
hormonal release: regulation by mental retardation proteins
Membrane traffic relies on three key mechanisms: the budding of vesicles starting from
a donor compartment, the displacement of these vesicles towards an acceptor
compartment, and their subsequent fusion. During regulated exocytosis, membrane
fusion involves the release of the vesicular contents in the extracellular medium. This
key mechanism allows the release of the neurotransmitters and the hormones. Fusion
requires the assembly of the SNAREs complex, which brings the vesicular membrane
closer to the plasma membrane, but also modifications of the lipidic composition of the
membranes at the exocytotic site. We recently discovered the crucial role of fusogenic

lipids in the release of the neurotransmitters and the hormones. It is now time to
address their dynamics and modes of synthesis. We will focus on phosphatidic acid (PA)
that is specifically produced at the exocytotic site by phospholipase D (PLD). Using PLD1
and PLD2 KO mice models, already in house, we propose to characterize physiological
parameters altered in these models, but also to look at individual fusion events using
electrophysiological approaches such as catecholamine release by chromaffin cells using
carbon fiber amperometry. On the other hand, we will use a targeted lipidomic approach
to characterize precisely the individual PA species that are produced at the exocytotic
sites and indentify those that have a specific role in membrane fusion. We will also study
precisely PA dynamics and turnover during exocytosis, with a focus on its potential
conversion in lysoPA and free fatty acids, of which we will estimate the potential
importance of poly-unsaturated forms.
Thanks to a model of KO mouse, we will also focus on the kinase RSK2 for which the loss
of function is responsible for the Coffin-Lowry syndrome, a pathology associating mental
retardation and skeletal defects. Indeed, we have just shown that RSK2 phosphorylate
PLD1 during exocytosis in chromaffin cells. This stage is required for the synthesis at the
exocytotic site of PA. From mice invalidated for RSK2, electrophysiological approaches
will be implemented in collaboration with a group in Bordeaux, in order to measure the
synaptic activity of the neurons in two complex circuits, obtained from culture in section
of cerebellum and of the amygdala. Measurements of the release of catecholamines by
amperometry, will also allow us to record the release of the stress hormones from
cultured mouse chromaffin cells. The dynamics of the interaction between RSK2 and
PLD1 will be studied by experiments of FRET, while the synthesis of PA will be studied
in real time thanks to a specific fluorescent probe for this lipid. Finally, the functional
importance of RSK2 in the synthesis and the dynamics of fusogenic lipids during the
release of hormones and the neurotransmitters will be compared with that of other
proteins involved in mental retardation.
1
/
2
100%