Titre : Organisation hiérarchique des connexions

Titre : Organisation hiérarchique des connexions corticothalamocorticales chez le chat.
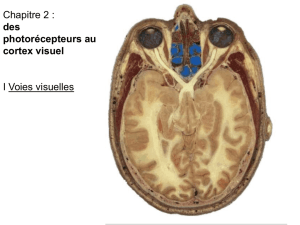
La perception visuelle émerge de l’encodage de caractéristiques optiques des stimuli dans
des voies fonctionnelles distinctes et de l’intégration progressive de l’information. Cette
intégration est effectuée dans des aires corticales qui sont organisées en réseaux
hiérarchiques. Cette hiérarchie est déterminée par la nature des connexions
corticocorticales feedback et feedforward (Rockland and Pandya 1979; DeYoe and
VanEssen 1988; Felleman and VanEssen 1991). Dans le système visuel, l’information du
cortex visuel primaire est transmise progressivement vers des aires corticales de niveaux
hiérarchiques supérieurs par des connexions feedforward corticocorticales.
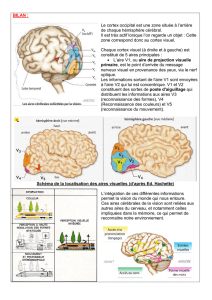
Plusieurs études récentes suggèrent que la communication entre les aires corticales ne
serait pas principalement assurée par des connexions corticocorticales mais par des circuits
cortico-thalamo-corticaux qui passent par des noyaux thalamiques de haut niveau
(Sherman and Guillery 1996; Guillery and Sherman 2002; Sherman and Guillery 2002;
Theyel et al. 2010). Le noyau latéral postérieur et le pulvinar sont parmi les noyaux
thalamiques de haut niveau les plus développés chez les primates. Ces noyaux reçoivent
des afférences de nombreuses aires corticales impliquées dans la vision et le contrôle
visuomoteur.
Il existe deux types de terminaisons axonales corticothalamiques. Les terminaisons de
petite taille avec une densité élevée de vésicules synaptiques qui contactent des dendrites
de petit calibre sont dites de type 1. Les terminaisons de plus grande taille, ayant une
moindre densité de vésicules synaptiques qui contactent des dendrites de plus grande
taille et qui forment des glomérules sont dit de type 2 (Mathers 1972a; Mathers 1972b;
Robson and Hall 1977a; Robson and Hall 1977b). Les terminaisons du type 2 prennent
origine des neurones de la couche V du cortex cérébral, et les terminaux du type 1
prennent origine des neurones de la couche VI (Bourassa and Deschenes 1995). Les
terminaux du type 1 forment des synapses dont les récepteurs postsynaptiques sont
principalement métabotropiques et, de ce fait, on les qualifie de terminaison ayant un rôle
modulateur. Les terminaisons du type 2 forment des synapses dont les récepteurs

postsynaptiques sont principalement ionotropiques et on les qualifie de terminaisons
« driver »(Sherman and Guillery 1998) .
La communication entre les aires corticales est donc établie par deux systèmes parallèles,
les connexions corticocorticales directes à partir desquelles on a défini la nature
hiérarchique du cortex cérébral ; et les connexions indirectes corticothalamocorticales. à
travers les noyaux thalamiques d’ordre supérieur.
Hypothèse Organisation hiérarchique des connexions corticothalamiques.
Puisque les synapses modulatrices du type 1 sont essentiellement des terminaisons des
axones des cellules de la couche VI et que les synapses driver du type 2 sont des
terminaisons des axones de la couche V, si les aires de bas niveau hiérarchique participent
davantage à des projections driver feedforward que les aires de haut niveau, on devrait
s’attendre à ce qu’un injection dans le pulvinar marque une plus grand nombre de
neurones de la couche 5 des aires de bas niveau hiérarchique et une plus grande
proportion de neurones dans la couche 6 des aires des niveaux hiérarchiques supérieurs.
Objectif :
L’objectif de cette recherche est de déterminer si les connexions corticocortical directe et
indirectes sont deux systèmes hiérarchiques parallèles. Ces travaux vont permettre de
connaître davantage le rôle des boucles cortico-thalamo-corticales dans la fonction visuelle
normale et de déterminer leur implication possible dans le maintien des fonctions visuelles
lors d’atteintes organiques.
Méthodologie
Des injections de traceurs neuronaux rétrogrades seront effectuées dans le noyau latéral
postérieur thalamique chez le chat et le cortex cérébral sera coupé en série de manière à
pouvoir quantifier la proportion de cellules marquées dans les couches V et VI des diverses
aires corticales.
Rôle de l’étudiant :
Assister et apprendre à effectuer des chirurgies stéréotaxiques et les injections dans les
noyaux thalamiques du chat. Apprendre et effectuer le traitement histologique du

matériel. Apprendre les éléments du logiciel Neurolucida nécessaires à effectuer des
cartographies des neurones maquées.
Période de stage :
Une période minimale de 8 semaine, de préférence 12 semaines :
Superviseurs de stage
Christian Casanova PhD
Professeur titulaire
École d’optométrie Université de Montréal

1.3 Bibliographie
References
Bourassa J and Deschenes M. 1995. Corticothalamic projections from the primary visual
cortex in rats: a single fiber study using biocytin as an anterograde tracer. Neuroscience.
66:253-263.
DeYoe EA and VanEssen DC. 1988. Concurrent processing streams in monkey visual
cortex. Trends Neurosci. 11:219-226.
Felleman DJ and VanEssen DC. 1991. Distributed hierarchical processing in the primate
cerebral cortex. Cereb Cortex. 1:1-47.
Guillery RW and Sherman SM. 2002. Thalamic relay functions and their role in
corticocortical communication: generalizations from the visual system. Neuron. 33:163-175.
Mathers LH. 1972a. The synaptic organization of the cortical projection to the pulvinar of
the squirrel monkey. J Comp Neurol. 146:43-60.
Mathers LH. 1972b. Ultrastructure of the pulvinar of the squirrel monkey. J Comp Neurol.
146:15-42.
Robson JA and Hall WC. 1977a. The organization of the pulvinar in the grey squirrel
(Sciurus carolinensis). I. Cytoarchitecture and connections. J Comp Neurol. 173:355-388.
Robson JA and Hall WC. 1977b. The organization of the pulvinar in the grey squirrel
(Sciurus carolinensis). II. Synaptic organization and comparisons with the dorsal lateral
geniculate nucleus. J Comp Neurol. 173:389-416.
Rockland KS and Pandya DN. 1979. Laminar origins and terminations of cortical
connections of the occipital lobe in the rhesus monkey. Brain Res. 179:3-20.
Sherman SM and Guillery RW. 1996. Functional organization of thalamocortical relays. J
Neurophysiol. 76:1367-1395.
Sherman SM and Guillery RW. 1998. On the actions that one nerve cell can have on
another: distinguishing "drivers" from "modulators". Proc Natl Acad Sci U S A. 95:7121-
7126.
Sherman SM and Guillery RW. 2002. The role of the thalamus in the flow of information to
the cortex. Philos Trans R Soc Lond B Biol Sci. 357:1695-1708.
Theyel BB, Llano DA and Sherman SM. 2010. The corticothalamocortical circuit drives
higher-order cortex in the mouse. Nat Neurosci. 13:84-88.

1
/
5
100%