BIOCHIMIE DE L`AXE HYPOTHALAMO

Page 1 sur 16
Endocrinologie : cours n°3
Jeudi 22 janvier 2009 de 10h30 à 12h30
Professeur : Pr DE ROUX
Ronéotypeur : Quentin FISCHER
BIOCHIMIE DE L’AXE
HYPOTHALAMO-HYPOPHYSAIRE

Page 2 sur 16
PLAN
I. Généralités
II. Anatomie fonctionnelle
a) Hypothalamus
b) Hypophyse
c) Système porte hypothalamo-hypophysaire
d) Organisation cellulaire de l’hypothalamus
e) Organisation cellulaire de l’hypophyse
f) Les axes endocriniens
III. Développement de l’hypophyse
IV. Biochimie du système hypothalamo-
hypophysaire
a) Neuropeptides
b) Récepteurs
c) Signaux intracellulaires
V. Les différents axes neuroendocriniens
a) Axe thyréotrope
b) Axe corticotrope
c) Axe somatotrope
d) Axe lactotrope
e) Axe gonadotrope
VI. Les neurones magnocellulaires

Page 3 sur 16
I. Généralités :
Le but de ce cours est de nous expliquer comment tout ce qu’on a appris en physiologie, en
biologie cellulaire et en biochimie peut être intégré dans un enseignement de science
médicale. Il est donc très important de connaître ces mécanismes pour comprendre la
physiopathologie moléculaire des maladies ainsi que la pharmacologie.
II. Anatomie fonctionnelle :
a) Hypothalamus :
L'hypothalamus est une structure indispensable à la vie, située dans le cerveau
(contrairement à l’hypophyse).
C’est un système intégrateur qui va recevoir, à la fois, des afférences neuronales,
notamment du cortex, des afférences endocriniennes, notamment de la périphérie
(venant des glandes endocrines) ainsi que des afférences métaboliques (exemple : la
glycémie donne des informations à l’hypothalamus qui répond par une augmentation
ou une diminution de la sensation de faim). Outre ces stimuli internes, on va avoir
également des stimuli externes avec notamment le rôle de l’environnement. Par
exemple, l’hypothalamus reçoit un signal directement de la notion nuit/jour. C’est
grâce à ce signal qu’on a la régulation du rythme circadien donc la régulation du
rythme biologique au cours du nycthémère.
Il participe au contrôle :
Des hormones périphériques (tout le système est basé sur des rétrocontrôles
qui régulent la production de neuropeptides au niveau de l’hypothalamus. Si
cet équilibre est perturbé, on aura des pathologies d’hyperfonctionnement ou
d’hypofonctionnement)
De la fonction motrice des organes digestifs
Du comportement alimentaire des individus
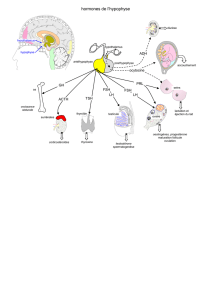
b) Hypophyse :
Elle est située dans la selle turcique (c’est donc une glande en dehors du cerveau)
Elle est composée de deux régions :
L’hypophyse antérieure qui est une glande endocrine
L’hypophyse postérieure appelée neurohypophyse car composée
d’extrémités axonales de neurones dont le corps cellulaire est situé dans
l’hypothalamus. Ces neurones sont les neurones magnocellulaires qui vont
synthétiser la vasopressine notamment.
Elle reçoit des afférences :
Hypothalamiques fondamentales
Périphériques qui peuvent être hormonales (ex : rôle des hormones
stéroïdes) ou métaboliques
Sa fonction est de synthétiser des peptides qui vont aller stimuler les glandes
endocrines périphériques, ainsi que le rein et l’utérus.

Page 4 sur 16
Du fait de la proximité du chiasma optique, on comprend que lors d’une expansion de
l’hypophyse, on peut avoir une compression du chiasma optique qu’il va falloir traiter
extrêmement rapidement pour éviter la perte de la vision.
La fonction neurosécrétrice passe
obligatoirement par l’éminence
médiane située à la base de
l’hypothalamus. C’est au niveau
de cette éminence médiane
qu’on a apparition du système
porte car la membrane hémato-
encéphalique est fenêtrée, ce qui
permet le lien entre
l’hypothalamus et l’hypophyse.

Page 5 sur 16
c) Système porte hypothalamo-hypophysaire :
Il y a des neurones dont les corps cellulaires sont
situés dans l’hypothalamus et dont l’extrémité
axonale est située dans l’hypophyse. Il s’agit de
la posthypophyse. Le lien se fait de façon
logique par l’intermédiaire des axones.
Pour l’antéhypophyse, il n’y a pas de lien
cellulaire direct : le lien passe par un réseau
vasculaire qu’on appelle le système porte qui a
donc pour fonction de recevoir des peptides
hypothalamiques et de les transporter à
l’hypophyse antérieure où ils vont pouvoir aller
activer leurs récepteurs spécifiques et donc
réguler la voie endocrinienne.
Ce système porte est très important et très développé chez l’homme, qui permet une
interaction étroite entre l’hypothalamus et l’hypophyse. Mais malheureusement, la plupart
des facteurs sécrétés par l’hypothalamus pour aller activer l’hypophyse antérieure ont des ½
vies très courtes, d’où une impossibilité de les doser. On peut faire par contre des études
pharmacologiques pour étudier la fonction de ses facteurs.
d) Organisation cellulaire de l’hypothalamus :
C’est une organisation très complexe car l’hypothalamus est composé de neurones,
neurosecréteurs ou non, et de cellules gliales de soutien. Ces neurones sont organisés en
noyaux. Leurs noms dérivent de leurs positions anatomiques ou de leurs formes selon la
position latérale, médiale ou périventriculaire et selon un axe antéropostérieur. Ces noyaux
possèdent une spécificité fonctionnelle et sont en inter-relation.
Il y a deux types de neurones :
Les neurones parvicellulaires qui ont une fonction neurosécrétrice vers le système
porte hypothalamo-hypophysaire. Ces neurones ont des corps cellulaires dans les
noyaux hypothalamiques et dont l’extrémité axonale est située au niveau de
l’éminence médiane. (l’éminence médiane est une structure qui ne possède pas de
barrière hémato-encéphalique c’est-à-dire qui a la possibilité de subir des
stimulations par des peptides dans la circulation sans pénétration dans le cerveau,
c’est la région qui permet l’interaction entre l’hypothalamus et le système vasculaire
dans le sens où c’est là que sont relargués tous les peptides de l’hypothalamus qui
sont synthétisés dans les neurones parvicellulaires)
Les neurones magnocellulaires qui ont également une fonction neurosécrétrice,
mais dans la veine hypophysaire. Ils synthétisent l’ocytocine et notamment la
vasopressine. La neurosécrétion va survenir dans la veine hypophysaire, c’est-à-dire
que l’extrémité axonale est située au niveau de la posthypophyse.
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%