Neurophysiologie Sensorialité

Neurophysiologie
Sensorialité
I. Généralités
Le fonctionnement du Système nerveuse se fait sur trois éléments : entrée/ intégration/ sortie.
Entrée : capter les informations de l’information a l aide récepteurs spécifiques qui
transforment les énergies environnementales en potentiel d’action> codage de l’information =
transduction. Puis par les voies afférentes les informations sont acheminés vers le SNC.
Intégration : analyse des informations par le SNC. Traitement rapide par la moelle épinière en
réflexe mais simultanément le SNC analyse pour créer une sensation et donc une perception.
> Ordre d’action partant par des voies efférentes vers le SNP.
Sortie : produire des comportements.
Tout le fonctionnement de l’organisme reproduit ce schéma.
1. Rôle des informations sensorielles :
Fonction d’alarme et de protection : il nous alerte sur la modification du milieu extérieur pour
permettre de modifié notre comportement.
Fonction d’analyse : analyser les informations de notre environnement et de les intégré dans
le temps et l’espace pour permettre l’exploration de notre environnement.
2. Classification des sensibilités :
Somesthésie : sensibilité générale :
Extéroceptive : superficielles cutanée
Nociceptive : sensibilité douloureuse
Proprioceptive : sensibilité musculaire
Intéroceptive : sensibilité des organes interne ;
Sensorialité : sensibilité desservie par des appareils sensoriels localisés :
Téléceptive : vision, audition, odorat > éloigné de l’organisme.
Chémoceptive : chimique (> le gout)
Labirinthique : équilibre
3. Classification des récepteurs sensoriels :
Mécanorécepteurs :
Sensibilité tactile cutanée : récepteurs tactiles (épiderme)
Sensibilité profonde : récepteurs des muscles et des tendons
Audition récepteurs chocléaire.
Equilibration : récepteurs vestibulaire
• Thermorécepteur : variation de température
• Nocicépeteur : douleur
• Récepteurs électromagnétique : vision
• Chémorécepteur : gout olfaction.
4. Organisation des 2 voies sensorielles :
• Voies spécifiques primaire constitué de trois neurones : organes des sens, motoneurones >
moelle épinière > bulbe >thalamus> air spécifique du cortex.

• Voies non spécifiques : elle pour départ la voies primaire, mais ce sont certain neurones qui
se sont séparés et se retrouve dans la formation réticulé > puis se retrouve dans des noyau non
spécifique du thalamus > qui vont activés l’ensemble du cortex. Cela à pour Rôle : augmenter
la vigilance de l’organisme, permet la prise de conscience de l’information et donc que nous
sommes capable de la reconnaitre et de l’identifier.
Elle détient des informations du système limbique (stimulus émotionnels) et de
l’hypothalamus (fonctions végétatives).
5. Organisation corticale : somatotopie
Ex la sensibilité de la peau :
PEAU :
Relais 1
Relais 2
Cortex
A travers chaque relais on trouvera une somatotopie > projection point par point.
LE TRAITEMENT DE L’INFORMATION SENSORIELLE :
Transduction :
Au niveau du récepteur il existe deux sites spécifiques :
- Site transducteur : transforme les stimuli sensoriels en énergie
- Site générateur : génère le PA
Un stimulus entraine, la dépolarisation de la membrane par l’entrée d’Ion Sodium (N+) > ce
qui déclenche un potentiel récepteur. Le site transducteur devient alors positif, ce qui entraine
alors un courrant électrique entre le site transducteur (+) et le site générateur (-). Cela entraine
un changement de polarité du site générateur, qui induit alors un potentiel générateur, qui
quand il atteint un certain seuil déclenche une émission de PA qui renferme toutes les
informations sensorielles.
Codage de l’information sensorielle dans le potentiel d’action :
1. La qualité (modalité sensorielle) > sensation éprouvée (visions audition, touché, goût,
odorat) et différente qualités de la sensation (vision : couleur/ audition : fréquence)
2. L’intensité : sensation forte ou faible.
3. La durée : sensation brève ou longue.
4. Localisation : localisation dans l’espace (visions, audition ou sur notre corps) et capacité à
différencier deux stimulus voisins.
Modalités sensorielles :
Organes sensoriels : les 5 sens > modalité sensorielle due à la nature du récepteur stimulé et
de la spécificité des voies sensorielles emprunté par l’information.

En effet chaque récepteur sensoriel possède des caractéristiques intrinsèques qui vont lui
permettre de capter un stimulus sensoriel.
Codage de l'information sensorielle-la qualité (modalité sensorielle) → qualités intrinsèques
du récepteur → spécificité des voies sensorielles
-l'intensité (1) → réponse à un stimulus faible donne naissance à un potentiel général → '' ''
modéré donne naissance à un potentiel générateur juste en dessous du seuil → réponse à un
stimulus fort donne naissance à un potentiel générateur supérieur au seuil ce qui donne le
potentiel d'action (PA)
Augmentation d'amplitude du potentiel générateur => création du Potentiel d'Action
Seuil absolu qui permet de mesurer la sensibilité d'un système sensoriel, plus l'intensité du
stimulus augmente plus la fréquence des PA est importante
(2)Lorsque le PA atteint 150 par secondes les neurones vont saturer (neurone à seuil bas,
neurone à seuil moyen, neurone à seuil élevé) Plus on augmente l'intensité plus ce sera des
seuils différents de seuil d'activation bas à seuil d'activation élevé
Le codage de l'intensité du stimulus se fait en trois étapes :
1- codage en amplitude du potentiel générateur
2- codage en fréquence des potentiels d'action
3- recrutement de neurones à seuil de plus en plus élevé
-la durée → stimulus de durée constante, émet des PA, la fréquence des PA va diminuer =
phénomène d'adaptation. Les récepteurs tactiles sont des récepteurs à adaptation rapide. Les
thermorécepteurs réagissent rapidement au variation de températures.
Les récepteurs articulaires et musculaires ne réagissent pas ou peu. Nocicepteurs.
-Récepteurs phasiques répondent au début de la stimulation puis cessent des répondre puis
reprennent la stimulation en fin. Rôle : codent les variations d''intensité du stimulus en
fonction du temps (aspect dynamique)
-Récepteurs toniques répondent pendant toute la durée de la stimulation. Rôle : valeur absolue
de la durée du stimulus (aspect statique)
Le codage de la durée du stimulus se fait grâce au mécanisme d'adaptation qui permet de
décrire deux types de récepteurs :
1- les récepteurs toniques à adaptation lente renseignent sur la durée du stimulus
2- les récepteurs phasiques à adaptation rapide renseignent sur les variations d'intensité du
stimulus dans le temps
-la localisation → détection Acuité → discrimination
Croisement des voies sensorielles Somatotopie permet d'affiner la localisation, mécanisme
relativement grossier Champ récepteur : tous les neurones sensoriels sont constitués par un
champ récepteur. C'est l'ensemble des récepteurs qui sont en relation avec ce neurone
sensoriel
Densité des récepteurs Taille du champ récepteurs peut varier : La précision de la localisation
et le pouvoir de discrimination dépendent de la taille des champs récepteurs
Corrélation entre le récepteur et la sensation et le récepteur et le neurone.

Phénomène d'inhibition latérale : Certains récepteurs sont en accès direct avec le neurone
sensoriel (centre), de chaque côté : périphérie récepteur en relation indirect par l'intermédiaire
de inter-neurone inhibiteur
Stimulus au centre va activer directement le neurone sensoriel donc émission de PA
centre : excitateur
périphérie : inhibitrice
La taille du champ récepteur peut varier. Stimulus étroit ou étendu
Grand champ récepteur et stimulus étroit → réponse peu activée
Petit champ récepteur et stimulus étroit → forte activité
Grand champ récepteur et stimulus étendu → forte activité
Petit champ récepteur et stimulus étendu → pas de changement d'activité
Seuil de discrimination tactile :Extrémité de la langue =/ dos
Effet de contraste Répartition spatiale du stimulus sans inhibition latéraleRépartition spatiale
du stimulus avec inhibition latéraleL'inhibition latérale améliore le contraste de l'information
II. Le système visuel
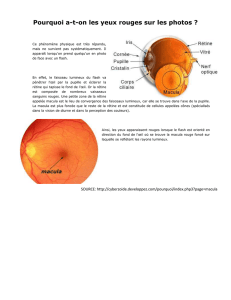
Il possède un organe spécialisé : l’œil
Il est constitué de différentes membranes superposées : sclérotique, la choroïde et la rétine
La rétine est constituée de deux sortes de récepteurs visuels: les bâtonnets et les cônes, qui
sont en relation avec des neurones bipolaires eux-mêmes en relation avec des neurones
ganglionnaires
Les neurones ganglionnaires forment de longs axones qui vont former le nerf optique
Entre les neurones bipolaires et les récepteurs visuels se trouve la cellule horizontale
L’information suit donc le chemin normal (récepteurs visuels > neurones bipolaires >
neurones ganglionnaires), ou passe directement par la cellule horizontale
Au niveau de la rétine, on trouve deux zones particulières : la fovéa (centre de la rétine) qui
est située sur l’axe optique ; puis la papille optique ou tâche aveugle (c’est le point de
convergence de tous les axones des cellules ganglionnaires)
La spécificité du récepteur rétinien est qu’il renferme une molécule de rhodopsine qui sert à
décomposer l’information
Organisation de la rétine
La densité des bâtonnets est importante à la périphérie de la rétine, proche du centre leur
nombre diminue rapidement jusqu’à ne plus en avoir au niveau de la fovéa
Les cônes sont très rares à l’extrémité de la rétine, augmentent lorsqu’on se rapproche du
centre et leur nombre devient maximum au niveau de la fovéa
Au niveau du point aveugle, il n’y a ni cônes ni bâtonnets
On distingue la rétine centrale et la rétine périphérique :
- La rétine centrale : à ce niveau, les relations entre les récepteurs, les cellules bipolaires
et ganglionnaires se font de manière linéaire
- La rétine périphérique : les relations entre les récepteurs et les cellules bipolaires se
font de manière convergente

Ces différences d’anatomie font que dans la rétine centrale, le champ récepteur du neurone
ganglionnaire est très petit car il est constitué par un seul un récepteur
Au niveau de la rétine périphérique, les champs seront plus étendus car les relations se font de
manière convergente
Rôles des cônes et des bâtonnets
Les bâtonnets sont des cellules ayant un seuil d’activation très bas, ils ont une forte
sensibilité à la lumière)
Ils sont utilisés en visions de nuit (vision scotopique)
Les cônes ont un seuil d’activation élevé donc ils sont peu sensible à la lumière
Ils sont donc plutôt utilisés en visions de jour (vision photopique)
Les cônes présentent cependant une particularité : ils sont responsables de la vision des
couleurs
Conséquences fonctionnelles :
Au niveau de la rétine centrale, un récepteur est en relation avec une cellule bipolaire, qui est
elle-même en relation avec une cellule ganglionnaire : les champs récepteurs sont donc très
petits
Au niveau de la rétine périphérique, une cellule ganglionnaire est en relation avec plusieurs
cellules bipolaires, qui sont elle-même en relation avec plusieurs récepteurs : les champs
récepteurs sont plus étendus
La rétine centrale va pouvoir détecter de manière précise la position d’un stimulus
La rétine périphérique quant à elle, va brouiller l’information
La précision de l’information sera moins grande au niveau de la rétine périphérique
La rétine centrale est plus précise que la rétine périphérique (acuité visuelle maximum)
Lorsqu’un stimulus est positionné sur la rétine centrale, un potentiel récepteur au niveau des
récepteurs va être transmis aux cellules bipolaires, puis aux cellules ganglionnaires
S’il est positionné sur la rétine périphérique, les potentiels récepteurs vont être transmis à
l’ensemble des récepteurs, puis aux cellules bipolaires et enfin aux cellules ganglionnaires
Un même stimulus de même intensité ne pourra être perçu par la rétine centrale, mais sera
perçu par la rétine périphérique
La rétine périphérique est plu sensible que la rétine centrale
Si l’on prend un stimulus en mouvement, il sera essentiellement détecté au niveau de la rétine
périphérique
La rétine périphérique est sensible aux mouvements
Propriétés fonctionnelles de la rétine
Rétine périphérique :
- Présence de bâtonnets
- Relations convergentes
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%