Ecologie, Systématique et Evolution - Université Paris-Sud

1
Muséum National
d’Histoire
Naturelle de Paris
Laboratoire des Reptiles et Amphibiens
25 rue Cuvier
75005 Paris
Impact de champs électromagnétiques sur la
faune protégée au large des côtes de Guyane
française
Par
Marc Girondot,
Professeur d’Ecologie et Environnement
à l’Université Paris Sud
Laboratoire d’Ecologie, Systématique et Evolution,
Université Paris-Sud et CNRS
Professeur au Muséum National d’Histoire Naturelle de Paris
Laboratoire des Reptiles et Amphibiens
Version du 10 janvier 2007
Rapport effectué à la demande de :
Parc Euromédecine 2
39 rue Jean Giroux
34080 Montpellier - France
Ecologie, Systématique et Evolution
Université Paris-Sud / CNRS / ENGREF - UMR C8079
Bât. 362 - 91405 Orsay cedex France
http://www.ese.u-psud.fr

2
1. Résumé
Les eaux de la Guyane française accueillent environ 26 espèces de cétacés et 5 tortues
marines qui sont toutes protégées en particulier
via
deux conventions internationales signées
par la France et deux protocoles. La France a en particulier obligation de prévenir une
modification significative et nuisible du milieu marin de la zone d'application de la Convention
et d’empêcher les espèces susmentionnées de voir leur statut se dégrader.
Une analyse détaillée de l’impact potentiel de méthodes utilisant un champ
électromagnétique pour la recherche des hydrocarbures a été effectué. Les champs électriques
sont reconnus par les requins mais il n’y a pas d’évidence qu’ils soient reconnus par les tortues
marines ou les cétacés.
Par contre le champ magnétique est clairement ressenti et utilisé par les tortues marines
et les cétacés et chez ces derniers des déviations aussi faibles que 13 nT pourraient être
ressenties. Un effet si faible est mesuré à 10 km de la source électromagnétique utilisée pour la
prospection. On notera cependant que les cétacés semblent plutôt être conduit vers la côte par
les zones de faible champ magnétique donc rajouter une composante ne créera pas de couloir
dans lesquels ils pourraient s’engouffrer. On ne doit pas donc s’attendre à avoir des échouages
de cétacés d’autant que cette prospection se passe fort loin de la côte.

3
Impact de champs électromagnétiques sur la
faune protégée au large des côtes de Guyane
française
2. Plan
1. Résumé...........................................................................................................................2
2. Plan................................................................................................................................3
2.1. Liste des figures ............................................................................................................................4
3. Introduction ...................................................................................................................5
3.1. Principe du SBL ...........................................................................................................................5
3.1.1. Qu’est-ce qu’un champ électrique ......................................................................................................... 7
3.1.2. Qu’est-ce qu’un champ magnétique ...................................................................................................... 8
3.1.3. Intensité d’un champ magnétique .......................................................................................................... 9
3.1.4. Le champ magnétique terrestre .............................................................................................................. 9
3.1.5. Le ferromagnétisme .............................................................................................................................. 12
3.2. Les espèces protégées dans les eaux de Guyane française ...................................................14
3.2.1. Les cétacés............................................................................................................................................. 14
3.2.2. Les tortues marines ............................................................................................................................... 16
4. Champs électromagnétiques et la faune.......................................................................16
4.1. Introduction ................................................................................................................................16
4.2. Les champs électriques et le vivant .........................................................................................16
4.2.1. La perception du champ électrique ...................................................................................................... 16
4.2.2. La pêche électrique ............................................................................................................................... 18
4.3. Les champs magnétiques et le vivant ......................................................................................19
4.3.1. La perception du champ magnétique ................................................................................................... 19
4.3.2. Les cétacés............................................................................................................................................. 21
4.3.3. Les chéloniens marins .......................................................................................................................... 24
4.3.4. Les chondrichtyens ............................................................................................................................... 26
5. Impacts environnementaux des champs électromagnétiques : synthèse ......................27
5.1. Caractéristiques de la source....................................................................................................27
5.2. Impacts pour la faune................................................................................................................27
6. Références bibliographiques ........................................................................................28

4
2.1. Liste des figures
Figure 1: Dépôt des récepteurs de champ électrique sur le sol................................................................................. 5
Figure 2: Le dipôle source situé à 40 m du fond transmet un signal électromagnétique répété à une fréquence
comprise entre 0,05 et 10 Hz. ....................................................................................................................................... 6
Figure 3: L'énergie électromagnétique est rapidement dissipée dans les sédiments conducteurs du fond, mais
l'est beaucoup moins dans des couches présentant une résistance plus forte, comme les réservoirs
d'hydrocarbures............................................................................................................................................................. 6
Figure 4: La réflexion de ce champ électromagnétique est enregistrée par les récepteurs posés sur le fond........ 7
Figure 5: A la fin, les récepteurs sont libérés du fond et remontent à la surface. Ils transmettent alors leurs
données à un satellite. ................................................................................................................................................... 7
Figure 6: Schéma montrant l'influence du champ magnétique terrestre................................................................... 9
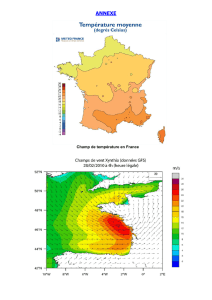
Figure 7: Tracé de l’intensité totale du champ magnétique terrestre en nT en 2005............................................. 10
Figure 8: (a) Schéma du champ magnétique terrestre montrant comment les lignes de champs (représentée par
des flèches) sont ressenties à la surface de la terre et comment l’inclinaison (l’angle formé entre la ligne de
champ et la surface de terre) varie avec la latitude. À l’équateur magnétique (la ligne courbée), les lignes de
champs sont parallèles à la surface de la terre et leur angle devient progressivement plus important que l’on se
rapproche des pôles où l’angle d’inclinaison est de 90°. (b) Schéma illustrant les 4 éléments du champ
magnétique terrestre qui peuvent, en principe, fournir aux animaux une information positionnelle. Le champ
magnétique en tout point peut être décrit en terme d’intensité totale ayant un angle d’inclinaison. Cette intensité
totale peut aussi être décomposée en deux vecteurs composants : une intensité horizontale et une intensité
verticale........................................................................................................................................................................ 11
Figure 9: Tracé de la composante horizontale du champ magnétique terrestre en nT en 2005............................ 11
Figure 10: Tracé de la composante verticale du champ magnétique terrestre en nT en 2005.............................. 11
Figure 11: Évolution prévue du champ magnétique terrestre total en nT/an de 2005 à 2010. ............................. 12
Figure 12: Acquisition d'un champ magnétique dans un matériau sous l'action d'un champ magnétique
extérieur. ...................................................................................................................................................................... 13
Figure 13: Exemple d'hystérésis sur un matériau dur. ............................................................................................. 13
Figure 14 : Distribution supposée des cétacés dans les eaux de Guyane française............................................... 15
Figure 15: Ampoule de Lorenzini chez un requin..................................................................................................... 17
Figure 16: Détection du champ électrique d'une proie potentielle par le requin Mustelus canis. ........................ 17
Figure 17: Détection du champ électrique par un jeune requin marteau. Les lignes rouges matérialisent les
lignes de champ électrique.......................................................................................................................................... 18
Figure 18: Impact de la dérive liée aux courants sur la capacité à trouver un point cible. .................................. 20
Figure 19: Carte des éléments stationnaires du ciel dans l'hémisphère nord......................................................... 21
Figure 20: Relation entre l'intensité du champ magnétique le long de la côte des USA et le nombre d'échouage
de cétacés. .................................................................................................................................................................... 22
Figure 21: Influence du champ magnétique local sur la probabilité d'échouage en fonction de l'espèce
(Kirschvink et al. 1986). La valeur en nT correspond à la déviation du champ magnétique expliquant le
maximum des échouages. Les espèces avec une relation significative échouage/ déviation du champ magnétique
sont montrées en gras.................................................................................................................................................. 23
Figure 22: Effet du champ magnétique sur l'orientation de jeunes Caouannes. .................................................... 25
Figure 23: Direction suivie par des tortues vertes juvéniles en fonction de leur lieu de nourrissage initial. ...... 26
Figure 24: Déviation du champ magnétique par rapport au champ magnétique terrestre en fonction de la
distance en x et y en mètre de la source..................................................................................................................... 27

5
Impact de champs électromagnétiques sur la
faune protégée au large des côtes de Guyane
française
3. Introduction
3.1. Principe du SBL
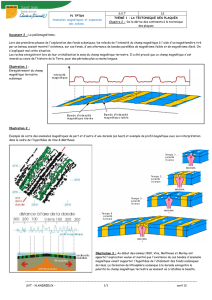
La technique de diagraphie des fonds marins prévue dans le cadre de cette étude est du
« seabed logging » (ou SBL). Il consiste à générer un champ électromagnétique à partir d’un
dipôle électrique horizontal mobile et à analyser sa réception grâce à des récepteurs de champ
électrique disposés sur le sol.
Les différentes phases sont présentées sur les Figure 1 à Figure 5.
Figure 1: Dépôt des récepteurs de champ électrique sur le sol.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
1
/
31
100%







![[4] Susceptibilités](http://s1.studylibfr.com/store/data/003629260_1-3ca03b480b86418dfcd84dc43138f11a-300x300.png)