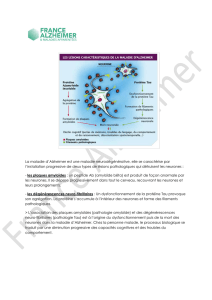
ÉTUDE DE L`ARBORISA`ITON AXONALE DE LA VOIE NIGROS
publicité

JULIE GAUTHIER ÉTUDE DE L'ARBORISA'ITON AXONALE DE LA VOIE NIGROS-E CaEZ LE RAT PAR MARQUAGE UMTAIRE Mémoire présen té à Ia Faculté des études supérieures de ITUniversitéLavai pour l'obtention du grade de mStre ès sciences (M-Sc.) Département d'anatomie et de physioIogie FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL Novembre 1998 O Julie Gauthier, 1998 National tibrary Bibliothèque nationale du Canada Acquisitions and Bibliographie Services Acquisitions et services bibliographiques 395 Wellington Street OttawaON K I A O N 4 Canada 395.rue Wellington OttawaON K1AON4 ChW?i The author has granted a nonexclusive licence allowing the National Libmy of Canada to reproduce, loan, distribute or sell copies of this thesis in microform, paper or eIectronic formats. L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la foxme de microfichelnlm, de reproduction sur papier ou sur format électronique. The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fiom it may be printed or otherwise reproduced without the author's permission. L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thése ni des extraits substantieis de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation. À mes parents REMERCIEMENTS Je remercie sincèrement mon directeur de recherche, le Docteur André Parent pour m'avoir donné la chance de faire ma maîtrise sous sa supervision et pour la confiance qu'il m'a démontrée. Son immense répertoire de connaissances scientifiques. sa grande disponibilité et sa sirnplicité m'ont permis de mieux orienter mes intérêts et de réaliser cet ouvrage. l e le remercie aussi pour m'avoir aidée à obtenir la prestigieuse bourse du Conseil de Recherches Médicales du Canada (CRM) qui a largement contribué à mon soutien financier durant toute la durée de mon étude. Je remercie également tous les membres de son équipe: Carole Émorid et Lisette Bertrand pour leur assistance technique et leurs conseils et les étudiants qui de près ou de loin ont participé à ma formation en tant que future chercheure. J'adresse un merci tout spécial à mes collègues et amis, Lucia Prensa (Ph-D.) et Ali Charara (Ph-D.), pour leurs précieux conseils, leur soutien scientifique et surtout pour leur support mord tout au long de ces deux années d'études au laboratoire. Finalement. je remercie tous les chercheurs du Centre de recherche Université Laval Robert-Giffard pour avoir partagé leurs connaissances scientifiques ainsi que Lucie Huart pour son support technique à la rédaction de ce mémoire. TABLE DES MATIÈEs Page REMERCIEMENTS ...................................................................................... ,,....................... IV ......................................................................................................... V TABLE DES MATIÈREs . LISTE DES ABREVMIIONS ........................,.,..................................................................... .VI LISTE DES FIGURES ..................., , . , ............................................................................... VlD ..................................................................................................... SUBSTANCE NOLRE 1 2.1 Substance noire pars cornpacta ............................................................................... 2 2.2 Substance noire pars réticulata ............................ . . . ............................................3 INTÉGRATLONDES NEURONES DOPAMINERGIQUES DANS LA CIRCUITERLE=DES GANGLIONS DE LA BASE . ................................. 4 3.1 Introduction aux ganglions de la base .................................................................... 4 .......................................................... 4 3.1.1 te striatum ....................................... . . , 3.1.2 le globus pallidus ............................................................................................ 7 3.1.3 le noyau sousthalamique............................................................................... 8 3.2 La projection nigrostriée ......................................................................................... 8 . ....................................................... 1 1 3.3 La projection nigropallidale ........................ 3.4 La projection nigrosousthalamique ...................................................................... 12 3.5 Fonctionnement des ganglions de la base ............................................................. 12 ..................................................................................... MALADIE DE PARKINSON 13 4.1 Physiopathologie ................................................................................................ 1 4 4.2 Affections neurochimiques ....... ........................................................................... -14 PROBLÉMATIQUEDE RECHERCHE 6.1 6.2 6.3 6.4 ...................................................................15 .............................................. 17 Animaux et préparation des tissus .................... . . Histochimie pour révéler le traceur .................................... .................................. 18 Immunohistochimie pour révéler la calbindineD28k ..........................................18 Analyse du matériel ..............................................................................................19 DISCUSSION ................................................................................................................ 27 BIBLIOGRAPHIE ................................................................................................................ -30 LISTE DES ABRÉVLAZIONS Noyau accumbens Commissure antérieure Acétylcholinestérase Biocytine Biotine Dextran Amine Carida1 CB: Calbindine-D28k, une protéine liant le calcium cc: Corps calleux Noyau caudé Cd: Ch: Striatum CTX: Cortex cérébral D: Dorsal Dopamine DA: Acide dihydroxyphénylacétique DOPAC: Dynorphine DYN: Ef: Noyau entopédonculaire Acide y-arnino-butyrique GABA: la décarboxylase, l'enzyme de 1' acide glutamique GAD: GB: Ganglions de la base GP: Globus pdlidus Globus pallidus, segment externe GPe: GPi: Globus pdlidus, segment interne HVA: Acide homovanif lique ic: Capsule interne Ventricule latéral LW M: Médian ML: Lemniscus médian MPTP: 1-rnéthyl4phényl- lT2,3,6-tétrahydropyndine NADPH-d: L'enzyme nicotinamide-adénine-dinucléotide phospha Noyau pédonculopontin PPN: Putamen PUT: R: rostral RRA: L'aire rétrorubrale ac: Acb: AChE: BIO: BDA: C: Rt: SN: SNc: SNc-d: SNc-v: SNI: SNr: STN: TH: VA: VL: VTA: ZI: Noyau réticulaire Substance noire Substance noire, pars compacta Substance noire, pars compacta, partie dorsale Substance noire pars compacta, partie ventrale Substance noire pars latéralis Substance noire, pars réticulata Noyau sousthdamique Thalamus Noyau thalamique ventral antérieur Noyau thalamique ventral latéral Aire tegmentaire ventrale Zone incerta LISTE DES FIGURES Fig. 1 Représentation des ganglions de la base dans le cerveau humain en vue frontale ......5 Fig. 2 Diagramme de la circuiterie des ganglions de la base dans des conditions normales..... 6 Fig. 3 Schéma des projections dopaminergiques mésostriées chez le rat en plan sagittal. ...10 Fig. 4 Schémas respectifs de ta circuiterie des ganglions de la base dans des conditions nomitle et parkinsonienne. .....................................................................................16 Fig. 5 Schéma du patron d'arborisation du premier type d'axone nigrostrié provenant d'un site d'injection de BDA au niveau de la substance noire pars compacta. ..............2 1 Fig. 6 Reconstruction bi-dimentionnelle d'un axone nigrostrié du premier type à partir ........22 d'un corps cellulaire laissant des collatérales dans le GP. .................... . . . Fig. 7 Reconstxuction, à partir de coupes sériées, d'un axone nigrostrié du premier type relié au corps cellulaire, en plan horizontal. ........................................................... 23 Fig. 8 Schéma du patron d'arborisation du deuxième type d'axone nigrostrié provenant d'un site d'injection de BDA au niveau de la substance noire pars compacta. ......24 Fig. 9 Photomicrographies de neurones de la SNc injectés à la BDA de façon extracelIuIaire et juxtacellulaire en coupe sagittale traitée pour la calbindine, ainsi que de terminaisons axonaies striatales et d'une dendrite épineuse. ............................................................. 2 5 Fig. 10 Photomicrographies de cellules marquées à la BDA illustrant 1'emplacement du neurone par rapport au tiers ventral et au tiers dorsal de la SNc et montrant les varicosités sur un axone nigral. ............................................................................- 2 6 La projection nigrostriée dopaminergique est reconnue depuis longtemps comme une voie stratégique dans la circuiterie des ganglions de la base. Le rôle important et critique de cette projection dans la fonction des ganglions de la base est illustré par les troubles moteurs observés dans la maladie de Parkins~nsuite à une dégénérescence de cette voie. Ainsi, les modèles animaux actuels de la maladie de Parkinson impliquent la destruction sélective des neurones doparninergiques de la substance noire. Cette projection nigrostriée innerve abondamment toute l'étendue du striahm et libère la dopamine qui représente I'un des importants neuromodulateurs modulant le flux de l'information à travers les structures faisant partie des ganglions de la base. Le présent travail tente de décrire en détail l'organisation anatomique de la projection nigrostriée chez le rat. Ces données s'inscrivent dans Ie cadre d'un projet global entrepris dans le laboratoire du Dr André Parent dont le but est de déchiffrer l'organisation anatomique et fonctionnelle des ganglions de la base. de mieux comprendre la signification des données obtenues dans ce travail, nous allons d'abord présenter une vue générale de I'organisation de la substance noire et des ganglions de la base, suivi d'un survol du phénomène que constitue la maladie de Parkinson- 2.0 LA SUBSTANCE NOIRE La substance noire (SN) est un des noyaux les plus volumineux du mésencéphale. EIIe forme un complexe avec deux autres structures adjacentes, I'aire teamentaire ventrale (VTA) et I'aire rétrorubrde (RRA), le tout renfermant la majorité des neurones doparninergiques du système nerveux centrd (Dahlstrom et Fuxe, 1964; Domesik et coll., 1983). Elle doit son nom à son contenu en pigments de neuromélanine qui lui confère une couleur noire lors d'une coupe fraîche du cerveau humain. Elle a fait l'objet d'un très grand nombre d'études depois la découverte de son implication dans la maladie de Parkinson. Elle est située directement au-dessus des pédoncules cérébraux, sur Ia base ventrale du tegmentum mésencéphalique et latéralement aux fibres du nerf oculomoteur. La SN des mammifères est divisée en deux parties principales: la substance noire pars compacta (SNc) et la substance noire pars réticulata (SNr). Une troisième partie, la substance noire pars latéralis (SNI) comprenant les fibres et les neurones situés dorsolatéralement et rostralement, a été décrite par certains auteurs (François et coll., 1984; Poirier et coll., 1983). Cette dernière partie ne sera pas considérée dans le texte qui suit. 2.1 La substance noire pars compacta (SNc) La SNc constitue la partie dorsale de la SN, formant un feuillet de cellules dopaminergiques (Bjorklund et Lindvall, 1984) couvrant la SNr. Elle correspond au groupe doparninergique A9 décrit par Dahlstrom et Fuxe (1964). La morphologie. le nombre et l'organisation des cellules de la SNc varient d'une espèce à une autre. Chez les primates, les neurones de la SNc forment une bande compacte de cellules situées le long de la bordure dorsale de la SNr et envahissent cette dernière à plusieurs endroits (Poirier et coll., 1983; Arsenault et coll., 1988). Chez les rongeurs, l'organisation de la SNc est beaucoup moins complexe. En effet, chez le rat, la SNc apparaît comme un mince feuillet honzontal, dense en cellules, et qui est facilement distinguable de la SNr (Poirier et coll., 1983). D'un point de vue cytologique, les neurones de la SNc sont de taïIles variées et de formes bipolaire ou multipolaire. Ils sont particulièrement remarquabIes par leurs dendrites qui s'orientent horizontalement et verticalement (Gulley et Wood, 1971; Juraska et coll., 1977; Halliday et Tork, 1986). Selon ces critères cytologiques, la SNc peut être subdivisée en une zone dorsale et une zone ventrale, La zone dorsale de Ia SNc se caractérise par la présence de cellules qui sont de petite taille, diffusément distribuées et qui possèdent des dendrites orientées principaiement suivant l'axe médiolatéral. Médialement, Ia zone dorsale de la SNc se fusionne imperceptiblement avec la VTA, une structure adjacente contenant des cellules de taille différente. Pour sa part, la zone ventrale de la SNc comprend un plus grand nombre de neurones volumineux dont les dendrites s'orientent dorsolatéralernent (Parent, 1996). En plus des critères morphologiques, des études irnmunohistochimiques ont démontré que les celluies de la SNc peuvent êtres divisées en deux sous-populations selon l'absence ou la présence de la caibindine, une protéine liant le calcium. Les cellules de la partie dorsale, communément appelée le tiers dorsal, sont riches en calbindine alors que les celluIes de la partie ventrale, connue sous le nom de tiers ventral, sont pauvres en calbindine (Gerfen et coll., l987a; Yamada et coll., 1990; Côté et coll., 1991). De plus, les pigments de neuromélanine, caractéristiques de la SNc des primates, sont plus abondants dans les neurones du tiers ventral que dans ceux du tiers dorsal (Gibb et Lees, 1991). Cette division tient compte aussi du fait que les neurones dopaminergiques de la SNc occupent une importance fonctionnelle du fait que les neurones de la partie ventrale sont beaucoup plus vulnérables que les neurones de la partie dorsale lors de certaines maiadies neurodégénératives, telle la maladie de Parkinson (Yamada et coll., 1990: Lavoie et Parent, 1991; Hirsch, 1992). La Snc est en grande partie formée de cellules dopaminergiques. Elle contient environ 10% de neurones qui ne sont pas dopaminergiques. Certaines études ont mentionné que les neurones dopaminergiques peuvent aussi renfermer d'autres substances teiles que la cholécystokinine (HoHelt et coll., 1980), la neurotensine (Hokfelt et coll., 1984a), l'enzyme décarboxylase de I'acide glutamique (GAD) (Ford et coll., 1995) et la calrétinine, une autre protéine liant le calcium (Rogers, 1992; Resibois et Rogers, 1992). La SNc contient aussi beaucoup de fibres irnmunoréactives pour la choline acétyltransférase, provenant du noyau pédonculopontin (PPN) situé caudalement à la SN. De plus, la SNc envoie des projections massives vers le noyau caudé (Cd) et Ie putamen (PUT), ce qui constitue la projection nigrostriée dont la dégénérescence est la principale cause de la maladie de Parkinson. D'autres projections doparninergiques atteignent le globus pallidus (GP) et le noyau sousthalamique (STN) (voir plus loin). Une autre caractéristique de la SNc est le fait qu'elle forme un continuum avec deux autres structures adjacentes, la VTA et la RRA, qui correspondent respectivement aux groupes dopaminergiques A10 et A8 décrits par DahIstr6m et Fuxe (1964). La VTA est située médialement par rapport à la SN. Elle est constituée de cellules globulaires, qui se distinguent de celles de la SNc par le fait qu'elles sont de plus petite taille (Poirier et coll., 1983; Halliday et Tork, 1986). De plus, leurs dendrites sont orientées horizontalement. La RRA est formée des cellules dopaminergiques situées caudalement par rapport à celles de la SNc et dispersées entre les fibres du lemniscus médian (ML). Les neurones de la RRA sont de forme multipolaire et de taille plus petite que ceux de la SNc. 2.2 La substance noire pars réticulata (SNr) La SNr est la partie de la SN qui se trouve ventralement par rapport à la SNc, tout juste audessus des pédoncules cérébraux. Comparativement à la SNc, elle contient peu de cellules qui se dispersent entre les fibres strïatonigrales et qui utilisent l'acide ramino-butyrique (GABA) comme neurotransmetteur (OerteI et coll., 1982; Smith et coll., 1989). Ces cellules GABAergiques de la SNr forment, avec celles du segment interne du GP (GPi), les portes de sortie des ganglions de la base. Ces neurones sont de taille moyenne ou grande et de forme ovale, ronde ou triangulaire (Juraska et coll., 1977; FaIlon et Loughlin, 1985). Ils possèdent des dendrites non-épineuses orientées rostrocauddement à l'exception de quelques neurones situés dans la partie ventrale de la SNr dont Ies dendrites sont parallèles aux pédoncules cérébraux (François et coll., 1987; Yelnik et coll., 1987). La SNr contient aussi beaucoup de fibres immunoréactives pour le GABA, la substance P, l'enképhaline, et la dynorphine (DYN) (Parent, 1996). De nombreuses évidences ont montré que ces fibres prennent leur origine des neurones du striatum (voir Parent, 1986). La SNr envoie des projections vers les noyaux thalamique ventral antérieur (VA) et ventral latéral (VL) et innerve aussi les collicules supérieurs et le PPN (Deniau et COIL, 1977; Graybiel, 1978; Parent, 1996). 3.0 INTÉGRATPONDES NEURONES DOPAMINERGIQCIES DANS LA ClRCtTIERIE DES GANGLIONS DE LA BASE 3.1 Introduction aux ganglions de la base Les ganglions de la base (GB) sont un ensemble de structures situées en dessous du cortex cérébral à la base du cerveau et impliquées dans le contrôle du comportement moteur et psychoaffectif. Ils sont constitués du striatum. du globus pallidus, du noyau sousthalamique et de la substance noire (Fig. 1). Ces structures sont connectées entre elles formant une circuitene distincte, étroitement reliée zu thalamus et au cortex cérébral (Fig. 2). Cette circuitene est dite extrapyramidale parce que les GB contrôlent la motricité mais que, contrairement aux fibres du système moteur corticospinal de type pyramidal, leurs axones ne passent pas dans les pyramides bulbaires. Dans le texte qui suit, nous allons présenter une vue générale de chacune des cornposantes des GB. à l'exception de la SN qui a fait l'objet du premier chapitre. 3.1.1 Le striatum Le striatum est la composante la plus volumineuse des GB. Chez les primates, le striatum est formé par le noyau caudé et le putamen qui sont séparés par la capsule interne, alors que, chez le rat, il apparaît comme une masse unique traversée par les fibres provenant du cortex cérébral, d'où le nom "putamen-caudé" chez cette dernière espèce. De plus, la partie rostroventrale du striatum, désignée comme striatum ventral, est formée par le noyau accumbens et le tubercule olfactif. Les études utilisant les techniques de Golgi et de microscopie électronique ont démontré que le striatum est constitué de deux groupes principaux de neurones que l'on peut distinguer d'après la présence ou l'absence d'épines sur leurs dendrites. Les neurones épineux, considérés comme des neurones de projection, sont de taille moyenne et représentent environ 95% de la population neuronale du striatum. Les neurones de projections peuvent être divisés en deux types selon leur contenu en neurotransmetteurs ainsi que leur cible préférentielle. Les Figure 1: Schéma d'une coupe d e cerveau humain en plan frontal illustrant l'emplacement des GB; le PUT, le Cd, le GPe, le GPi, le STN et la SN. À noter que la SN, cette structure noire de forme ovoïde, se situe dans la partie inférieure du mésencéphale à la limite du tronc cérébral. Coupe frontale de cerveau humain Figure 2: Diagramme de la circuiterie des GB dans des conditions normales, Les connexions inhibitrices sont représentées par des flèches noires et les connexions excitatrices par des fieches blanches. Circuiterie des GB dans des conditions normales Tronc cérébral MoëIie épinière neurones qui projettent au GPi et la SNr contiennent du GABA, de la substance P et de la DYN. Les neurones projetant au segment externe du GP (GPe) referment du GABA et de I'enképhaline (Fonnum et coll., 1978; Reiner et Anderson, 1990). Dans la population des neurones nonépineux, considérés comme des intemeurones, on retrouve des neurones de taille moyenne et des neurones géants. Ils ne constituent qu'environ 5% de la population neuronale du striatum. On peut différencier quatre types d'intemeurones selon leur phénotype chimique. Tous les neurones moyens sont GABAergiques. Cependant on peut les classer en trois types selon la colocalisation avec d'autres substances chimiques. On retrouve des interneurones contenant de la parvdbumine, d'autres renfermant de la calrétinine, et des interneurones contenant à la fois de la somatostatine, le neuropeptide Y et l'enzyme nicotinamide-adénine-dinucléotide phosphatediaphorase (NADPH-d) (Cowan et coll., 1990; Kita et coll., 1990; Smith et Parent, 1986; Difiglia et Aronin, 1982). Les interneurones géants sont cholinergiques (Difiglia et coll., 1976; Difiglia, 1987; Parent, 1986, 1996; Parent et coll., 1995). La distribution de plusieurs marqueurs neurochimiques permet aussi de subdiviser le striaN m en deux compartiments, soit les striosomes et la matrice extrastriosomde. Les striosomes correspondent à des îlots pauvres en calbindine et en acétylcholinestérase (AChE), l'enzyme de dégradation de l'acétylcholine. Pour sa part la matrice extrastriosomale, représentant 80% du volume striatal, est riche en cdbindine et en AChE (Graybiel et Ragsdale, 1978; Gerfen, 1985). Cette organisation neurochimique a une importance fonctionnelle au niveau des GB puisque l'organisation des connexions striatales suit aussi ce patron striosomes/matrice (Graybiel, 1990; Gerfen, 1992). Le striatum est considéré comme la porte d'entrée des GB puisqu'il reçoit des afférences massives de presque toutes les régions du cortex cérébrai, mais aussi de la SNc, de la VTA, de la RRA, des noyaux intralaminaires thalamiques, et des noyaux raphé dorsal et médian. D'un autre côté, le striatum projette massivement au niveau des deux segments du GP ainsi que de la SNr- En plus de ces connections extrastriatdes, il existe des connections intrinsèques propres au striatum (voir revues par Parent, 1996; Parent et Hazrati, 1995) 3.1.2 Le globus pdlidus (GP) Le GP est situé médialement par rapport au putamen dont il est séparé par Ia lame médullaire externe. Ii est divisé à son tour chez les primates en deux segments: soit un segment externe (GPe) et un segment interne (GPi), séparés par la lame médullaire interne. Chez les rongeurs, le GP, l'homologue du GPe des primates, se situe dorsolatérdement, tandis que le noyau entopédonculaire (EP), I'homologue du GPi des primates, se trouve ventromédialement par rapport au GP et il est entouré par les fibres de la capsule interne (Parent, 1996). Comme le Cd et le PUT, les deux segments du GP forment une seule entité cytologique contenant essentiellement des neurones de forme ovale ou polygonale avec de longues dendrites lisses et épaisses (Fox et cotl., 1974). Les cellules pallidales utilisent le GABA comme neurotransmetteur inhibiteur (Oertel et coll,, 1984; Smith et coll-, 1987). Le GP établit des connections réciproques avec le striatum et le STN. Les projections pallidosousùialamiques et pallidostriatales prennent naissance dans le GPe, tandis que les projections sousthalamopallidales se terminent à la fois dans le GPi et le GPe. Les neurones du GPi projettent massivement vers le thalamus formant ainsi l'une des principales portes de sortie des GB. Les projections pallidothalamiques, cheminent le long de I'anse lenticulaire et du faisceau lenticulaire pour se terminer dans les noyaux VA, VL et le noyau centromédian du thalamus. La projection pallidothalamique donne en plus des collatérales se terminant dans PPN et I'habénuta. En plus de ces connections, le GP envoie des projections vers la SNr, principalement à partir du GPe (voir revues Parent et Hazrati 1995; Parent, 1996). 3-1.3 Le noyau sousthaIamique (STN) Le STN est une petite structure en forme d'amande qui, comme le nom l'indique, est situé en dessous du thalamus, à la Iimite entre le prosencéphale et le tronc cérébral- Le STN est constitué d'une variété de celluIes dont la majorité sont de grande taille ou de taille moyenne et de formes pyramidale ou ronde avec des dendrites relativement longues et plus ou moins épineuses (Rafols et Fox, 1976; Paxinos, 1985; Parent, 1996). Une petite population cellulaire fusiforme a été décrite en marge du STN (Parent, 1996). Le STN utilise le glutamate comme neurotransmetteur excitateur (Smith et Parent, 1988). Il reçoit différentes afférentes surtout en provenance du GPe, du cortex cérébral et du PPN. Le STN à son tour projette vers les deux segments du GP ainsi que vers la SNr (Parent, 1996; Parent et Hazrati, 1995). 3.2 La projection nigrostriée L'existence de la projection nigrostriée a été rapportée, il y a très longtemps, suite aux observations de dégénérescence rétrograde des neurones au niveau de la SN successivement à d'importantes lésions du stnatum (Mettler, 1943). Par la suite, l'utilisation de la technique d'histofluorescence pour la visualisation des monoamines (Falck et coll., 1962) a permis de décrire la présence de trois groupes de cellules dopaminergiques désignés A8, A9 et A10, dont les axones atteignent le striatum @ahIstrom et Fuxe, 1964; Andén et coll., 1966; Unprstedt, 197 1). Plus tard, l'utilisation des méthodes de traceurs rétrograde et antérograde a confirmé l'existence de la projection nigrostriée et a permis d'élucider certains aspects de la topographie de cette projection. De plus, les études immunohistochimiques à l'aide d'anticorps dirigés contre la dopamine, ou contre la tyrosine hydroxylase, l'enzyme limitante dans la synthèse des catécholamines, ont permis de décrire l'organisation de cette projection ascendante (Hokfelt et coll., 1984b; Arsenault et coll-, 1988; Lavoie et coll., 1989). II est connu présentement que la voie nigrostriée innerve l'ensemble du territoire striatd et prend son origine principalement au niveau de la SNc et, à moindre degré, dans la RRA. Les neurones de la VTA contribuent à la formation du système méso-limbo-cortical, dont les fibres se terminent dans la partie ventrale du striaturn et aussi dans le cortex frontal (Bjorklund et Lindvall, 1984; Hokfelt et coll., 1984b) (Fig.3). Les axones de ces trois stnicnires dopaminergiques s'accumulent dorsornédialement par rapport à la partie rostrale de la SN. as traversent le champ "H" de Fore1 et montent le long du faisceau prosencéphalique médian circulant dans l'aire hypothalamique latérale pour atteindre plusieurs des structures des GB, principalement le striatum (Lavoie et coll., 1989). La projection nigrostriée est surtout ipsilatérale. les fibres controIatéraIes n'excédant pas 5% du nombre total de fibres de cette projection (voir Parent, 1996). Cette projection semble suivre une organisation topographique chez différentes espèces. En effet, chez le singe, la partie rostrale de la SNc projette au Cd, la partie caudale projette au PUT alors que les neurones de la VTA projettent principalement au striatum ventral (Szabo, 1980; Carmona et coll., 1991). D'autres études utilisant des traceurs rétrogrades fluorescents chez le singe ont montré que les cellules de la SNc projetant au PUT, et celles qui projettent au Cd forment des îIots distribués tout le long de la SNc sous forme d'une mosaique (Parent et coll., 1983). Chez le rat, Gerfen et ses collaborateurs ont montré que la projection nigrostriée est organisée en deux systèmes selon les projections vers les deux compartiments du striatum, les striosomes et la matrice extrastriosomaie (Gerfen, 1987a, 1987b). Les cellules de la VTA, de la RRA et du tiers dorsal de la SNc projettent vers la matrice et expriment I'imrnunoréactivité pour la calbindine, tandis que les neurones de la partie ventrale de la SNc et ceux qui forment des petits îlots dans la SNr projettent vers les striosomes et ne sont pas immunoréactifs pour la calbindine (Gerfen et coll., 1987a, 1987b). Cependant, les données irnmunohistochimiques ont montré que les fibres dopaminergiques innervent d'une façon uniforme tout le territoire du striatum, bien que les sûiosomes soient moins densément innervés comparativement à la matrice (Lavoie et coll., 1989). Figure 3: Projections dopaminergiques nigrostriées chez le rat en vue sagittale. Les neurones doparninergiques de la SNc innervent principalement le striaturn dorsal tandis que le striamm ventral, comprenant entre autres le noyau accumbens (ac), reçoit les projections doparninergiques de la VTA. Coupe sagittale de cerveau de mt Des observations en microscopie électronique ont montré que les fibres dopaminergiques nigrostriées forment des contacts synaptiques de type symétrique principalement avec la base des épines dendritiques des neurones de projectioîia dii striatum (Freund et coll., 1984). Ainsi les terminaisons dopaminergiques sont bien placées pour pouvoir moduler l'influence des fibres corticostriatales qui se terminent principalement sur la tête des épines dendritiques des cellules striatales, 3.3 La projection nigropallidale La présence des fibres catécholarninergiques au niveau du GP fut d'abord observée chez l'embryon humain et ensuite chez le rat par Bjorklund et collaborateurs (Nobin et Bjorklund, 1973; Lindvall et Bjorklund, 1979) en utilisant la technique d'histofluorescence pour la visualisation des monoamines. L'analyse détaillée chez le rat a amené Lindvall et Bjorklund (1979) à conclure que l'innervation catécholaminergique du GP provient des collatérales de la projection nigrostriée et que ces fibres forment des contacts intimes avec les ceIIules palliddes. Par la suite, les études imrnunohistochimiques chez le singe utilisant la tyrosine hydroxylase comme marqueur des neurones dopaminergiques ont démontré l'existence de projections dopaminergiques vers le GP, projections dont les axones s'arborisent principalement dans le GPi (Lavoie et coll., 1989). Chez le singe, cependant, des études effectuées à l'aide de méthodes de double marquage rétrograde combinées à l'immunohistochirnie pour la tyrosine hydroxylase suggèrent que la voie nigropallidale aurait une origine cellulaire distincte de celle de la projection nigrostriée (Smith et al. 1989). Récemment, l'injection d'un traceur rétrograde dans le GP chez le singe a montré que les neurones marqués sont distribués d'une façon uniforme dans la SN avec une légère prédominance à la jonction entre la SNc et la VTA (Charara et Parent, 1994). Les observations préliminaires en microscopie électronique indiquent que les fibres dopaminergiques forment des synapses de type "en passant" avec les cellules du GP chez le singe (Parent et Lavoie, 1993). Tout récemment, une étude effectuée chez le rat a rapporté que les fibres immunoréactives pour la tyrosine hydroxylase montrent des arborisations variqueuses ainsi que des boutons très espacés, qui occasionnelIement forment des contacts intimes avec les cellules cholinergiques qui se trouvent dans le GP (Rodrigo et coli., 1998). Ces fibres forment principalement des contacts synaptiques symétriques avec les dendrites cholinergiques (Rodrigo et coll., 1998). La fonction de la projection nigropallidaie reste encore mai connue. Les expériences électrophysiologiques ont montré que la dopamine peut produire une augmentation de L'activité des cellules pallidales chez le rat et atténuer l'effet inhibiteur du GABA injecté iontophorétiquement (Bergstrom et Wdters, 1984; Nakanishi et coll., 1985). Par ailleurs, les études chez les singes rendus parkinsoniens suite à l'injection de la neurotoxine 1-méthyl-4-phényl-1,3,3,6tétrahydropyndine (MPTP), ont démontré que la projection nigropailidale est relativement épargnée comparativement à la projection nigrostriée dans ce modèle animal de la maladie de Parkinson (Parent et coll., 1990; Schneider et Dako, 1991). L'ensemble de ces travaux laissent suggérer un rôle important de la dopamine au niveau du GP. 3.4 La projection nigrosousthalamique Le STN est une structure entourée de plusieurs fibres doparninergiques prenant origine au niveau de la SNc, la VTA et la RRA. La présence des varkositées axonales catéchoIarninergiques au niveau du STN fut d'abord détectée par les études d'histofluorescence chez le rat (Bjorklund et Lindvall, 1984), le chat (Meibach et Katzman, 1979) et l'humain (Nobin et Bjorklund, 1973). L'injection de traceurs rétrogrades dans le STN produit aussi du marquage rétrograde des cellules au nivea.; de la SNc chez le rat (Campbell et coll., 1985), le chat (Rinvik et coll., 1979) et le singe cynornolgus (Rinvik et coll., 1979). Cependant, une étude irnmunohistochimique effectuée chez le singe écureuil, a rapporté que le STN ne reçoit pas une dense innervation doparninergique (Lavoie et coll., 1989). Tout récemment, l'utilisation de la méthode de traçage rétrograde en combinaison avec I'immunohistochirnie pour la tyrosine hydroxylase chez le rat, a confirmé l'existence d'une projection nigrosousthalamique dopaminergique (Hassani et coll., 1997) qui prend naissance dans la SNc et la VTA. Bien que la fonction de cette projection est inconnue, le fait que le STN occupe une position stratégique dans la circuiterie des GB, laisse supposer qu'elle peut exercer un rôle important en modulant l'activité du STN. 3.5 Fonctionnement des ganglions de la base Suite à des observations anatomique, électrophysiologique et clinique, Albin et coll. (1989) ont proposé un modèle de l'organisation fonctionnelle des GB. Ce modèle porte essentiellement sur la stimulation ou l'inhibition de deux voies, l'une directe et l'autre indirecte: A. La voie "directe" prend origine des cellules striatales contenant du GABA, de la substance P et de la DYN et projettant aux GPi et la SNr. L'activation de cette voie résulte en une inhibition du GPi et de la SNr. Ces deux structures donnent origine à une autre projection GABAergique innervant les neurones excitateurs du thalamus qui projettent en retour au cortex cérébral. Ainsi une inhibition du GPi résulte en une désinhibition du thalamus facilitant ainsi les mouvements initiés par le cortex cérébral. B. La voie "indirecte" prend origine des cellules striatales contenant du GABA et de l'enképhaline et projettant vers le GPe. Du GPe, une projection GABAergique innerve le STN qui à son tour envoie une projection excitatrice vers le GPi et la SNr. L'activation de la voie indirecte augmente l'activité inhibitrice du GPi et de la SNr sur les neurones du thalamus, ce qui a pour effet de réduire le mouvement initié par le cortex cérébral. La projection doparninergique nigrostriée contrôle ces deux voies en stimulant la première et en inhibant la deuxième. Parmi les récepteurs dopaminergiques que l'on retrouve dans le cerveau, ce sont les sous-types DI et D2 qui sont les plus abondants au nivcau du striatum, cornparativement aux autres sous-types (D3, D4et DS). Les récepteurs Dl sont surtout exprimés par les neurones GABAergiques contenant la substance P et la DYN, tandis que les récepteurs D,- sont surtout exprimés par les neurones GABAergiques contenant l'enképhaline. Ainsi, I'activation de la projection nigrostriée libère la dopamine qui, par action sur les récepteurs DI, stimule la voie directe et, en agissant sur les récepteurs D2, inhibe la voie indirecte. Bien que couramment ütilisé en clinique, ce modèle montre des failles, particulièrement en ce qui a trait à l'organisation anatomique des voies directe et indirecte. Un autre point faible du modèle est le fait qu'il néglige la possibilité d'une action de la dopamine ailleurs qu'au niveau des neurones stnataux (Parent et Cicchetti, 1998). Cependant, les données anatomiques démontrant l'existence des projections nigropallidales et nigrosousthalarniques suggèrent un rôle modulateur important de la dopamine non seulement au niveau du striatum mais aussi au niveau du GP et du STN- 4.0 MALADIE DE PAREUNSON James Parkinson fut le premier, en 1917, à décrire les symptômes de la maladie, qui porte son nom (Agid et al, 1987). Cette maladie neurodégénérative est l'une des plus fréquentes touchant 1% de la population âgée de 65 ans et plus. Cette affection est répandue sur toute la surface de la terre et présente peu de variations régionales. Les gens atteints de cette maladie présentent des symptômes cliniques tels que le tremblement des membres au repos, de l'akinésie, de la rigidité et des troubles de la marche et de la posture. Les patients présentent aussi des affections au niveau du système nerveux périphérique, engendrant différents troubles végétatifs, tels des problèmes digestifs, sexuels et de sommeil. On ne connaît pas encore la véritable cause de cette pathologie bien que plusieurs hypothèses soient actuellement considérées. L'excitotoxicité ainsi que la présence de radicaux libres dans le milieu extracellulaire au niveau de la SNc viennent en liste des hypothèses les plus plausibles. Des toxines environnementales, tels le MPTP, les pesticides et la présence de virus ainsi que l'influence de facteurs génétiques sont d'autres causes possibles de la maladie de Parkinson (Olanow, 1987). 4.1 Physiopathologie La dégénérescence des neurones doparninergiques pigmentés à I'origine de la voie nigrostriée et, par conséquent, la baisse de dopamine au niveau striatal est la caractéristique biochimique la plus importante de la maladie de Parkinson (Agid et al, 1987; Parent, 1996). Les cellules du locus coeruleus et parfois même celles du noyau basalis de Meynert sont aussi affectées. Dans 84 à 100% des cas, les neurones en dégénérescence contiennent des inclusions éosinophiles appelées corps de Lewy (Barbeau, 1986). Ils sont constitués de composantes du cytosquelette, tels les neurotubules. Le pourcentage de mort ceIlulaire au niveau de la SN varie de 50 à 85% et ce sont les cellules calbindine-positives qui sont le plus épargnées, c'est-à-dire les neurones se situant dans Ie tiers dorsal de la SNc et la VTA. Le système nigrostrié est plus sévèrement affecté que le système méso-limbo-cortical, ce qui implique que le niveau de dopamine est moins affecté au niveau du striatum ventral que du striatum dorsal. Cependant le PUT est en général plus sévèrement atteint que le Cd (Parent, 1986). Les symptômes de la maladie apparaissent Iorsque près de 80% des neurones sont dégénérés. La dégénérescence cellulaire de la SNc apporte des changements chimiques au niveau du striatum, comme la baisse de la dopamine, de l'acide dihydroxyphénylacétique @OPAC, métabolite de la dopamine), de l'acide homovanillique (HVA) et de La tyrosine hydroxylase (Agid et coll., 1987). Les niveaux de noradrénaline, sérotonine et substance P sont aussi diminués. Par contre, le GABA et le peptide Méthionine-enképhaline augmentent (Olanow, 1987). 4.2 Affections neurochimiques On sait aujourd'hui que la dopamine agit au niveau striatal principalement via deux récepteurs, soit les sous-types DI et le Dî. Selon le modèle dudistique courant, impliquant une voie striatofuge directe et une voie indirecte (Gerfen, 1995; Wichman et DeLong, 1996), la transmission synaptique dopaminergique agirait de deux façons. Elle exercerait une action excitatrice sur le comportement moteur via les neurones striataux qui sont à l'origine de la voie directe et qui expriment le récepteur Dl- En revanche, elle exercerait une action inhibitrice en agissant sur les neurones striataux à I'origine de la voie indirecte et qui expriment le récepteur dopaminergique DZ.La figure 4 nous présente l'organisation générale de la circuiterie et I'activité neuronale des GB dans des conditions normale et pathologique (maladie de Parkinson). Une perte des cellules dopaminergiques de la SNc affecte toute cette circuiterie. Les voies de sortie, soit le GPi et ia SNr, reçoivent moins d'inhibition de la part des neurones stnataux. Il y a donc une augmentation de l'inhibition vers le thalamus ce qui entraîne une importante baisse d'activité des neurones thaiamocorticaux d'où l'apparition de l'akinésie ou de ta bradykinésie, des mouvements moteurs anormaux. En plus de cet effet médié par la voie directe, la perte de dopamine entraîne une désinhibition du STN via la voie indirecte. Cette désinhibition du STN permet à ce dernier d'augmenter son effet excitateur sur les neurones GABAergiques des voies de sortie des GB. Le tout résulte en une plus forte inhibition des neurones prémoteurs, thalamocorticaux, ce qui se traduit par une diminution de l'activité motrice. 5.0 PROBLÉMATIQUEDE RECHERCHE Jusqu'à maintenant aucune recherche ne s'est intéressée à la principale voie dopamineigique du système nerveux central à I'échelle unitaire. Avec une nouvelle technique expérimentale récemment développée dans notre laboratoire, on a voulu élucider l'organisation anatomique de la projection nigrostriée en portant une attention particulière à I'arborisation axonaie de cette projection. La technique d'injection cellulaire utilisée permet de marquer un seul neurone ou une très petite population de neurones identifiés de façon éIectrophysiologique. Cette méthode assure une visualisation détaiIlée du corps cellulaire et du patron d'arborisation axonale. Le marquage neuronal unitaire à été combiné avec I'immunohistochimie pour vérifier s'il existe une relation entre le patron d'arborisation d'un axone au niveau du striatum et les compartiments suiataux, soit les strïosomes et la matrice. De plus, en utilisant la même approche expérimentale, on a voulu vérifier s'il existait des différences dans le patron d'arborisation axonale entre les cellules du tiers ventral et du tiers dorsal de la SNc. Figure 4: Diagramme de la circuiterie des GB dans des conditions normale (diagramme de gauche) et pathologique (maladie de Parkinson), (diagramme de droite). L' activité neuronale normale des noyaux est représentée en ,gis moyen, une augmentation de l'activité est ombragée gris foncé et une baisse de I'activité est ombragée gris pâle. Les connexions inhibitrices sont représentées par des flèches noires, les connexions excitatrices par des flèches blanches. 6.1 Animaux et préparation des tissus Un total de 38 rats (Sprague-Dawley), de poids variant entre 250 et 300 p,ont été utilisés pour cette étude. Les procédures d'anesthésie et de chirurgie ainsi que les conditions d'hébegement des animaux de l'animalerie du Centre de recherche Université Laval Robert-Giffard (CRULRG) répondent aux normes établies par le Conseil canadien de protection des animaux. De plus, notre protocole expérimental a été approuvé par le comité de L'Université Laval sur l'éthique et l'utilisation d'animaux de laboratoire. Les animaux ont d'abord été anesthésiés par une injection intramusculaire ou péritonéale d'un mélange d'hydrochlorure de kétamine (Ketaset, 75 mgkg) et de xylazine (5 mgkg) et placés ensuite dans un appareil stéréotaxique. Ils ont reçu des injections microiontophorétiques bilatérales, parfois multiples, d'un traceur antére grade soit la biocytine (BIO. N-biotynil-L-lysine; Simgna,St. Louis, MO) soit La biotine dextran amine (BDA; Sigma) dans différents secteurs de la SNc, utilisant les coordonnées stéréotaxiques de l'atlas de Paxinos et Watson (1986). Cette procédure d'injection permet d'obtenir une visualisation détaillée et complète de I'arborisation axonde des neurones marqués. Brièvement, toutes les injections ont été faites sous guidage électrophysiologique avec des rnicropipettes de verre d'un diamètre variant entre 3 et 6 Fm. Les micropipettes étaient remplies d'une solution d'acétate de potassium ( 0 5 M ) avec 2% de BI0 ou de BDA. Dans tous les cas, un appareil iontophorétique de haut voltage ((Neurodata) a été utilisé afin d'injecter le traceur en appliquant un courant positif de 300 à 400 nA (1 sec O N / l sec OFF). Pour définir la nature des neurones dopaminergiques nous nous sommes servi des caractéristiques électrophysiologiques des neurones de la SN afin de choisir le site d'injection. La micropipette était d'abord descendue dans la SN afin de pouvoir détecter le patron de décharge typique des neurones de la SNr. Ces neurones montrent une activité très rapide et tonique (Grace et Bunney, 1984a,b), ce qui permet de les détecter plus facilement que ceux de la SNc (Grace et Bunney, 1984a,b). Ces deniers ont un patron de décharge lent et irrégulier, parfois en bouffée, mais le plus souvent ces neurones demeurent relativement silencieux, donc difficiles à détecter. Suite à l'identification électrophysiologique des neurones de la SNr et grâce aux coordonnées de l'atlas de Paxinos et Watson (Paxinos et Watson, 1986), la micropipette était légèrement remontée jusqu'au niveau de la SNc. Les animaux ont été sacrifiés après un temps de survie nécessaire pour le transport axonal soit de 24 heures dans le cas de Ia B I 0 et 72 heures dans le cas de la BDA. Ils ont été anesthésiés avec le mélange d'hydrochlomre de kétamine et de la xylazine et ensuite perfusés. Après ouverture de la cage thoracique, l'aorte fut canulée et l'animal fut perfusé avec une solution composée de paraformaldéhyde (4%) et de glutaraldéhyde (0.1%) dans du tampon phos- phate (PB, 0.1 M, pH 7 4 , et finalement par une solution à sucrosé 10% dans de PB (0.1 M, pH 7.4). Une fois prélevé, les cerveaux ont été post-fixés dans du sucrose 30% pendant une nuit à K et coupés à l'aide d'un microtome à congélation en sections de 60 F m d'épaisseur dans le plan sagittai ou horizontal. Toutes les sections sont recuillies dans un tampon phosphate salin (PBS, OJM, pH 7.4).rincées plusieurs fois et traitées pour révéler le traceur et aussi, dans la majorité des cas, la calbindine. 6.2 Histochirnie pour révéler le traceur Toutes les sections ont été traitées pour révéler la B I 0 selon le protocole décrit par Honkawa et Arsmstrong (1988) ou la BDA (Lévesque et coII., 1996)- Les sections ont été incubées pendant une nuit à 4 C dans une solution contenant le complexe avidine-biotine-peroxydase (ABC, Vector Labs, Burlingame, CA) dilué 1:100 dans du PBS (0.1 M, pH 7.4) et 1% Triton X-100 . La peroxydase a été révélée en incubant les sections dans une solution contenant du 33'-tétrahydrochlorure de diarninobenzidine (O.Os%, DAB ;Si-ma), du nickel d'ammonium suIfate(0.6%) (Fisher Scientifique) et du peroxyde d'hydrogène H,02 - (0.003%) dans du tarnpon Tris (0.05M, pH 7.6) pendant 10 à 15 minutes à la température de la pièce. La réaction fut arrêtée avec un rinçage dans du tampon Tris et deux rinçage dans un tampon PBS. Le DAB intensifié au nickel est un chromogène qui produit une coloration bleu foncée dans les éléments marqués, ce qui les rend très faciles à détecter- 6 3 Immunohistochimie pour révéler la calbindine-D28k Dans la plupart des cas, après avoir révélé le traceur, les sections ont été traitées suivant un protocole imrnunohistochirnique permettant de visualiser la cdbindine. Elles ont été incubées avec un anticorps anti-calbindine fait chez la souris (Sigma) dilué 1:2500 dans une solution de PBS contenant 1% de sérum normal de cheval (NHS) et 0-1% de Triton X-100 pendant une nuit à 4 C Le lendemain, les sections ont été rincées trois fois dans du PBS et incubées dans une solution de PBS contenant l'anticorps secondaire biotynilé (fabriqué chez le cheval et dirigé contre les IgG de la souris; Vector Labs, Burlingame, CA; dilution 1:200), 1% de NHS et 0.1% de Triton X100 pendant une heure à la température de la pièce. Après trois rinçages dans du PBS, les sections ont été incubées dans une solution ABC (dilution 1: 100 dans du PBS) pendant une heure à la température de la pièce selon le protocole décrit par Hsu et al. (1981). Le DAB a été utilisé comme chromogène, conférant aux éléments marqués une couleur brunâtre et uniforme comparativement au chromogène NiDAB qui donne une coloration grandaire bleu foncé. Les sections ont été ensuite rincées une fois dans du Tris et deux fois dans du PBS pendant 5 à 10 minutes à la température de la pièce pour arrêter la réaction. Toutes les sections ont été étalées sur des lames enrobées de gélatine, déshydratées et recouvertes de lamelles avec du Permount afin d'être examinées en microscopie optique. 6.4 Analyse du matériel La localisation et l'étendue précise des sites d'injection, pour chacun des rats utilisés dans la présente étude, ont été examinées avec un microscope optique de type Leica. Certains cas ont été exclus de l'analyse car les sites d'injection n'étaient pas situés au niveau de la SNc. Les dessins de I'arbonsation entière des axones niagaux individuels ont été faits à partir de sections sériées qui ont été analysées à l'aide d'une caméra lucida, à des grossissements de 20 à 40 x. Les champs d'arborisation des axones marqués ont été reconstruits à faibIe grossissement dans le but de déterminer leur localisation dans les GB. Des 38 rats ayant reçu des injections de tracews antéropdes au niveau de la SNc, seulement 24 avaient des sites d'injections qui étaient strictement confinés à la SNc. Chez les 14 autres mimaux Ies injections étaient situées dans la SNr, la VTA ou encore trop dorsalement ou trop ventraiement par rapport à la SNc. Les deux traceurs ont donné d'excellents résultats, mais la BDA a fourni un marquage plus foncé de l'axone des neurones de la SNc que la BIO. En revanche, la B I 0 donnait une image plus claire du domaine somatodendritique des neurones de la SNc. Nous avons donc utilisé les deux traceurs de façon complémentaire. En tout, quinze axones ont pu être entièrement reconstruits. Certains ont été tracés depuis le corps cellulaire d'origine jusqu'aw terminaisons striatales. Dans ces cas, les neurones d'origine étaient localisés dans chaque partie de la SNc, soit le tiers ventral et le tiers dorsal. D'autres axones ont été dessinés à partir du site d'injection, les neurones d'origine n'ayant pu être identifiés à cause du précipité bleu foncé qui empêche de voir les corps cellulaire. Cependant, l'analyse en profondeur des coupes adjacentes antérieures et postérieures au site d'injection a confirmé que ces axones émergeaient bien de la SNc. De plus, les quelques axones doparninergiques nigrostriés pouvant provenir du tronc cérébral, caudalement à la SNc Fig. 3), tels que certains axones émergeant du noyau raphé dorsal, ne passent pas par la SN, mais se concentrent et circulent plutôt dans la VTA, latéralement à l'hypothalamus (Paxinos, 1985). Les corps cellulaires des neurones de la SNc marqués avec le traceur étaient tous de taille moyenne ou de grande taille et donnaient origine à plusieurs dendrites longues, lisses et pauvrement arborisées. Dans quelques cas, on retrouvait des épines réparties uniformément sur les dendrites des neurones de la SNc (Fig. 10C). Les dendrites des cellules injectées, situées parmi les neurones calbindine-négatifs du tiers ventral de la SNc (Fig. 9A, Fig. lOA), étaient pour la plupart orientées dorsoventralement, tandis que les dendrites des cellules calbindine-positives localisées dans le tiers dorsal de la SNc étaient principalement alignées Ie long de l'axe médiolaîéral (Fig. 9B). La majorité des axones marqués émergeaient de dendrites p h a i r e s (Fig. 6B)et certains émettaient des collatérales axonales locales. Les axones obserués étaient très minces et pourvus de varicosités eparties à intervalles réguliers (Fig. 10B). Certains de ces axones bifurquaient caudalement en quittant la dendrite primaire d'origine, pour ensuite revenir dans la capsule interne et se rendre jusqu'au striatum. De plus, tous les axones étudiés présentaient une boucle axonale au départ, peu après avoir quitté le corps ceIldaire (Figs. 6-8). L'étude des coupes sériées du matériel injecté nous révèle l'existence d'au moins deux types d'axones nigrostrïés. L'axone du premier type (lY15) (Figs. 5-7) atteint le striatum directement en traversant la capsule interne et le GP. Au niveau du striaturn, cet axone s'arborise fréquemment et son champ d'arborisation temiinal couvre une région striatale relativement grande. Parmi ces fibres de premier type, quelques-unes émettaient une ou deux collatérales courtes au niveau du GP (Figs. 6 et 7). Une caractéristique importante du premier type d'axone était la présence de deux champs d'arborisations distincts au niveau du siriaturn (Fig. 7). Un premier champ d'arborisation très massif se situait près de la bordure du GP et l'autre, beaucoup plus petit que Ie premier et relié à celuici par une fine collatérale, se localisait dans la partie plus rostrolatérde du striaturn. Les deux champs d'arborisation se retrouvaient approximativement sur le même pIan dorsoventral. Le champ terminal des axones du type 1 est composé d'amas fibreux compacts de forme ronde ou ovale dispersés à travers une grande portion du striatum (Fig. 5B). À l'intérieur de chaque amas, des terminaisons formant un bulbe en apposition avec les neurones striataux ont été souvent observées. Contrairement au premier type d'axone, les axones du deuxième type (3/15) (Fig. 8) s'arborisent relativement peu au niveau du striatum. En revanche, ces axones fournissent de multiples collatérales à différentes composantes des GB, incluant I'EP, le GP et le STN. D'autres collatérales ont aussi été vues au niveau de la portion ventrale de la zone incerta où iI y a fusion avec ie noyau réticulaire thalamique et dans certains noyaux intralaminaires rostraux du thalamus. Sur des sections préparées pour la visualisation de la cdbindine, on a vu des axones traverser Ies striosomes du striaîum, mais I'arborisation occupait surtout la matrice extrastriosomale. Cette observation est valable dans les deux types d'axones révélés dans cette étude. Étant donné le faible nombre d'axones analysés ici, il ne nous a pas été possible de savoir s'il y avait une localisation préférentielle (tiers dorsaVtiers ventral) des neurones de type 1 et de type 2. Cette dificulté vient en partie du fait que quelques-uns des axones que nous avons tracés émergeaient d'un site d'injection où il n'était pas possible de localiser le corps cellulaire d'origine. De plus, certains neurones marqués étaient situés à la limite des tiers ventral et dorsal de la SNc. Figure 5: Représentation schématique dans le plan sagittal du patron d'arborisation d'un axone nigrostrié du premier type reconstruit à partir de coupes sériées. Cet axone émerge d'un petit site d'injection à la BDA (point rouge) situé dans la SNc chez le rat. L'axone parcourt la capsule interne (ic), le GP et s'arborise profusément dans Ie striatum (Ch).Le dessin en A donne une vue d'ensemble de cet axone en coupe sagittale. L'encadré B donne une vue plus détaillée de l'arborisation de l'axone dans le Ch. Figure 6: Représentation schématique dans le plan sagittal du patron d'arborisation d'un autre axone nigrostrié de type I reconstruit à partir de coupes sériées. Cet axone prend origine d'un seul neurone marqué à la BDA situé dans le tiers ventral de la SNc et s'arborise dans le pôle rostrd du striatum (CPu). Le dessin en A donne une vue d'ensemble en plan sagittal du chemin parcouru par l'axone marqué. Le dessin en B montre le détail des dendrites et du corps cellulaire. L'axone principal quittant une dendrite primaire est représenté en bleu pâle. C : Agrandissement d'une partie du champ terminal au niveau striatd. Figure 7: A: Reconstruction d'un axone nigrostrié à partir de coupes sériées de rat rnontruit. en plan horizontal, I'arborisation axonaie d'un axone de type 1. L'axone est connecté P un corps ceIlutaire localisé dans la SNc juste en bordure de la SNr. On remarque les deux champs d'arborisation au niveau du striatum. Un champ apparaît très massif près du GP et l'autre moins important plus rostralement. La pointe de flèche nous indique la région qu'on voit en plus forte magnification en B. B: L'axone pnncipai du neurone nigrostrié est représenté en bleu pâle et ses multiples collatérales sont en rouge. Figure 8: Représentation schématique sur le plan sagittal du patron d'arborisation d'un axone nigrostrié du deuxième type originant d'un petit site d'injection de BDA (point rouge) au niveau de la SNc reconstruit à partir de coupes sagittales sériées. Ce neurone était situé plus caudalement que celui illustré dans la Fig. 8A. L'axone s'arborise dans le striatum (CPu), le GP, le noyau entopédonculaire (EP), le noyau réticulaire (Rt), de même que dans le thalamus (TH). Le schéma en A nous présente une vue d'ensemble en plan sagittal du chemin parcouru par l'axone injecté. Les figures B et C montrent I'arborisation axonale détaillée au niveau du GP, de I'EP et du Rt (pointe de flèche), respectivement. Figure 9: A: Photomicrographies montrant deux neurones de la SNc (pointe de flèches) injectés de façon extracellulaire à la BDA au niveau du tiers ventral de la SNc. À noter le neuropile irnmunoréactif pour la calbindine qui caractérise la SNr, mais qui est absent dans la SNc. B: Un neurone de ia SNc marqué après injection juxtaceilulaire de BDA dans le tiers dorsal de la SNc. C: Partie d'une dendrite pourvue d'épines appartenant à la cellule injectée montrée en B. D: Arborisation dense au niveau du striatum d'un seul axone marqué à la BDA appartenant à un neurone de la SNc du premier type (voir Figs. 5.7). E: Arborisation striatale diffuse d'un seul axone marqué à la BDA d'un neurone de la SNc du deuxième type (voir Fig. 6). La barre étalon en A=150 pin ( aussi vaiide pour B), celle qui est en C=10 pm et en D=100 prn (aussi vaiide pour E). Figure 10: Photomicrographies montrant différents aspects de neurones de la SNc injectés par de la BDA. A: Un corps cellulaire dans la partie dorsale de la SNc. À noter Ies corps cellulaires positifs pour la caibindine dans la partie dorsale de la SNc, ainsi que le neuropile riche en calbindine au niveau de La SNr. B: Un autre neurone marqué à la BDA situé dans la partie dorsale de la SNc. À noter les nombreuses varicosités sur l'axone émergeant du neurone (flèche). 8.0 DISCUSSION L'utilisation d'une nouvelle technique d'injection jiuttacellulaire et extracellulaire de traceurs antérogrades biotynilés développée récemment dans notre laboratoire, nous a permis pour la première fois de visualiser des axones de la projection nigrosuiée à I'échelle unitaire chez le rat. Cette méthode puissante nous a permis de définir le patron d'arborisation de certains axones faisant partie de la principale projection doparninergique de la circuiterie des GB impliqués dans la dégénérescence neuronale associée à La maladie de Parkinson. Pour la première fois, nous démontrons, contrairement au schème actuel, que la voie nigrostriée n'est pas une entité monolithique. En effet, notre étude a révélé que ce système majeur de projection est composé d'au moins deux sous-systèmes anatomiquement distincts. Ces deux sous-systèmes sont caractérisés par les deux types de fibres nigrofûges observées dans la présente étude. Un des aspects frappants de l'organisation de cette projection est son patron d'arborisation relativement focalisé au niveau du striatum. Cette caractéristique de la projection nigrostriée va à l'encontre de ce que l'on croyait précédemment, à savoir que les axones nigrostriés s'arborisaient sur presque toute I'étendue rostrocaudale du striahm (Dalstrom et Fuxe, 1964; Fallon et Loughlin, 1985). De plus, le patron d'arborisation des axones nigrostriés, diffère remarquablement de celui des projections corticostrÏataIes et thalamostriatales tel qu'observé antérieurement avec la même approche expérimentale que celle utilisée dans la présente étude. En effet, le marquage unitaire des neurones corticostriataux et thdarnostriataux a révélé une arborisation striatale plus étendue que celle des axones nigrostriés (Lévesque et coll., 1996; Parent et coll., 1996). Des comparaisons intéressantes peuvent êtres faites avec nos présentes données chez le rat et les résultats antérieurs obtenus chez les singes normaux et les singes rendus parkinsoniens suite à l'administration de la neurotoxine MPTP (Parent et coll., 1990). Ces études chez Ie singe ont démontré que les fibres dopaminergiques s'arborisant dans le GP sont relativement épargnées chez le singe parkinsonien, contrairement à la perte massive de fibres observées au niveau striatal. Quelques fibres doparninergiques étaient néanmoins présentes dans le striatum de tous les singes MPTP examinés. Cette faible arborisation axonale au niveau du striatum présente des ressemblances remarquables avec celle des neurones de la SNc du rat qui branchent profusément dans les structures extrastriatales mais pauvrement dans le striatum (type 2). En effet, ces axones striataux demeurés intacts chez le singe parkinsonien étaient peu nombreux et branchaient très peu. Ces résultats suggèrent la possibilité que les neurones de type 2 pourraient être moins susceptibles à la dégénérescence dans la maladie de Parkinson que les neurones qui s'arborisent profusément dans le striatum (type 1). À cet égard, il serait très intéressant de savoir si les neurones de type 2 sont préférentiellement localisés dans le tiers dorsal de la SNc chez le rat. Cette région de la SNc renferme la majorité des neurones dopaminergiques qui expriment la caibindine (Gerfen, 1992) et qui sont relativement épargnés chez le singe parkinsonien (Lavoie et Parent, 1991). Au contraire, les neurones doparninergiques du tiers ventral qui sont dépourvus de cette enzyme dégénèrent massivement dans la maladie de Parkinson (Yamada et coll., 1990; Lavoie et Parent, 1991). L'hypothèse voulant que les neurones de type 2 soient relativement épargnés alors que les neurones de type L soient plus spécifiquement affectés serait en accord avec l'idée que les mécanismes neurotoxiques du iMFTP agiraient principalement au niveau du stria(Langston, 1985; McCrodden et coll., 1990). Ainsi, la susceptibilité des neurones dopaminegiques à I'action du MFTP pourrait être proportionnelle au degré d'arborisation de leurs axones au niveau striatal. Il est couramment admis que les neurones du tiers ventral de la SNc projettent aux striosomes, tandis que ceux du tiers dorsal s'arbonsent dans la matrice extrasû-iosornale (Gerfen, 1992). Cependant, les deux types d'axones nigrostriés identifiés dans la présente étude ne semblent pas respecter cette cornpartimentalisation striosomes/matrice. Bizn que nos données reposent sur un échantillon relativement petit. d'autres observations mettent aussi en cause la corrélation itablie par Gerfen (1992) entre la Iocalisation des neurones de la SNc e t leur patron d'innervation striatale. Entre autres, cette corrélation explique mal le fait que les nombreux neurones du tiers ventrai de la SNc, sont ceux que l'on dit innerver spécifiquement les striosomes, qui ne représentent que 20% du volume total du striatum (Lanca et coll., 1986). Dans cette perspective, il est aussi difficile d'expliquer la dénervation doparninergique massive qui implique l'ensemble du striatum, suite à la dégénérescence des neurones du tiers ventral observés dans la maladie de Parkinson. Nos données actuelles peuvent être comparées en partie à d'autres données intéressantes obtenues antérieurement à l'aide de la méthode d'histofluorescence à l'acide glyoxilique sur !'innervation doparninergique du GP chez le rat. Lindvall et BjorkIund ont suggéré en 1979 qu'il y avait une innervation dopaminergique du GP provenant probablement des collatérales de neurones de la SNc se rendant au striatum. Par contre, des études effectuées à l'aide de traceurs rétrogrades chez le singe ont conclu à l'existence d'une voie nigopallidale directe (Smith et coll ., 1989). Notre technique actuelle d'injection extracellulaire confirme 1' innervation doparninergique du GP par les neurones nigrostriés. De plus, elle démontre qu'un même axone nigrostrié laisse des collatérales dans d'autres composantes des GB, tel I'EP et le STN. Des études pharmacologiques et neurochirniques antérieures avaient suggérés I'existence d'une innervation dopaminergique au niveau du STN (Campbell et coll., 1985; Hassani et COL,1997; Meibach et Katzman, 1979),sauf qu'on n'avait pas démontré que cette innervation prenait naissance de collatérales d'un seul axone nigrostrié. Nos résultats nous indiquent clairement que l'organisation de la voie nigrostriée est plus cornpiexe que ce que l'on croyait. Bien que ces résultats soient préliminaires, on peut tout de même tirer une règle générale concernant l'organisation de la voie nigrostriée suite à cette étude. La voie nigrostriée est constituée d'au moins deux types de fibres avec deux patrons d'arborisation différents, ce qui permet un effet direct et spécifique via le premier type d'axones et un effet plus d i f i s sur les composantes extrastriataies via le second type. Une étude beaucoup plus poussée de ce système nous permettrait de savoir si Ie type d'axone arborisant dans plusieurs structures des GB (type 2) a un site préférentiel d'arborisation comme c'est le cas pour les neurones süiatofuges (Lévesque et coll., 1996;Parent et coll., 1996). Ces deux types de neurones pourraient jouer un rôle crucial (et complémentaire) dans l'organisation fonctionnelle des GB. Les neurones de type 1 s'arborisant profusément au niveau du striatum exercemient un effet massif et spécifique sur les neurones striataux, mais agiraient indirectement sur les voies de sortie des GB (EP/SNr). À l'inverse, les neurones de type 2 s 'arborisant principalement au niveau des structures extrastriatales, exerceraient une action plus faible sur les neurones striataux, mais agiraient directement sur les voies de sortie des GB. On peut imaginer que ces deux types d'axones fonctionnent de façon complémentaire dans des conditions normales. Cependant, dans une situation pathologique, comme la maladie de Parkinson, la synergie entre ces deux types de projections pourrait être sérieusement altérée. En conclusion, l'ensemble de nos observations nous permet de suggérer l'existence d'une règle fondamentale qui préside à l'organisation de la voie nigrostriée. Cette règle repose sur les deux types d'axones décrits dans la présente étude. Elle implique qu'un axone nigrostrié s'arborisant massivement dans le sûiatum innerve peu les structures extrastriatales, alors qu'un axone qui s'arborise profusément dans les structures extrastriatales, innerve peu le striatum. Afin de valider cette règle, il faudra bien sûr effectuer d'autres études visant à reconstruire un plus grand nombre d'axones nigrostriés. La taille de l'échantillon dépendra du nombre de type d'organisation axonale observé durant l'étude. Ce genre d'études nous permettrons de savoir s'il existe une distribution préférentielle des neurones de type 1 et de type 2 dans la SNc et d'établir la relation entre ces neurones et la compartimentaiisation striataie de type striosome/matrice. Agid, Y., Javoy-Agid, F. and Ruber, M. (1987). Biochemistry of neurotransrnitten in Parkinson's disease. Movernent Disorders, 2: 166-230, Albin, RL., Young, A.B. and Penney, J.B. (1989). The functionai anatorny of basal ganglia disorders . Trends Neurosci., 12:366-375. Andén, N.-E., Dahlstrom, A., Fuxe, K., Larsson, K., Olson, L. and Ungerstedt, U. (1966). Ascending monoamine neurons to the telencephalon and diencephalon. Acta Physiol. Scand., 67:3 13-326. Arsenault, M.-Y., Parent, A., Séguéla, P. and Descarries, L. (1988). Distribution and morphological characteristics of dopamine-immunoreactive neurons in the midbrain of the squirrel monkey (Saimiri sciureus). J. Comp. Neurol., 267:489-506. Barbeau, A. ( 1986). Parkinson's disease: clinical features and etiopathology. Handbook of CZinical Neurology, vol5 (49): 87-152. Bergstrom. D A . and Walters, J.R. (1984). Dopamine attenuates the effects of GABA on single unit activity in the globus pallidus. Brain Res., 3 10:23-33. Bjorklund, A. and Lindvail, 0.(1984). Dopamine-containing systems in the CNS. In A. Bjorklund and T. HokfeIt (Eds.). Handbook of Chernical Neuroanatomy. Classical Neurotransmitters in the CNS. Vol. 2, Part 1, Elsevier, Amsterdam, pp. 55-1 12. Campbell, GA., Eckardt, M J. and Weight, F.F. (1985). Dopaminergic mechanisms in su bthalamic nucleus of rat: Analysis using horseradish peroxydase and rnicroiontophoresis. Brain Res., 333 :261-270. Carmona, A., Catalina-Herrera, CJ. and Jimenez-Castellanos, J. (199 1). Nigrocaudate and nigroputaminal projections in the monkey. Acta mat., 141: 145- 150. Charara, A. and Parent, A. (1994). Brainstem dopaminegic, cholinergie and serotoninergic afferents to the pallidum in the squirrel monkey. Brain Res., 640:155- 170. Côté. P.-Y., Sadikot, A.F. and Parent, A. (1991). CompIementary distribution of cal bindin D-28k and parvalbumin in the basal forebrain and midbrain of the squirrel monkey. Eur. J. Neurosci., 3:1316-1329. Cowan, RL., Wilson, CJ., Emson, ' C . and Heizmann, C.W. (1990). Paravalbumin-containing GABAergic interneurons in the rat neostriatum. J . Cump. Neurol., 302: 19'7-205. Dahlstrom, A. and Fuxe, K. (19CX). Evidence for the existence of monoamine-containing neurons in the central nervous system. 1. Demonstration of monoamines in the ce11 bodies and brainstem neurones. Acfa Physiol- Scand (suppl .), 232: 1-55. Deniau, J.M., Harnrnond, C. Féger, J. and Mckenzie, J.S. (1977). Bilateral projection of nigrocolicular neurons: An electrophysiological analysis in the rat. Neurosci. Lett., 545-50. DiFiglia, M. (1987). Stinus organisation of cholinergic neurons in the monkey neostriatum. J. Comp. Neurol., 255:245-258. DiFiglia, M. and Aronin, N. (1982). Ultrastructure features of immunoreactive somatostatin neurons in the rat caudate nucleus. J. Neurosci., 2: 1267-1274. DiFiglia, M., Pasik, P. and Pasik, T. (1976). A Golgi study of neuronal types in the neostriatum of monkeys. Brain Res., 114:245-256. Domesik, V.B., Stinus, L. and Paskevitch. P.A. (1983). The cytology of dopaminergic and non-dopaminergic neurons in the substantia nigra and ventral tegmental area of the rat: A light and electron microscopic study. Neuroscience, 8:743-765. Falck, B., Hillarp, NA.,Thieme, G . and Torp, A- (1962). Fluorescence of catecholamines and related cornpounds condensed with formaldehyde. J. Histochem., 10:348-354. Fdlon, J.H. and Loughlin, S.E. (1985). Substantia nigra. In: Paxinos, G., The Raf Nervous System (2nd Ed), vol. 1, Academic Press, San Diego. Fonnum, F.,Gottesfeld, 2. and Grofova, 1. (1978). Distribution of glutamate decarboxylase, choline acetyltransferase and aromatic amino acid decarboxylase in the basal ganglia of normai and operated rats. Evidence for sûiatopallidal, striato endopeduncular and striatonigral GABAergic fibers. Brain Res., 143:125- 13 8. Ford, B., Holmes, CJ., MainvilLe, L. and Jones, B.E.(1995). GABAergic neurons in the rat pontornesencephalic tegmentum: Codistribution with cholinergic and other tegmental neurons projecting to the posterior lateral hypothalamus. J. Comp. Neurol., 363: 177-96. Fox, C.A., Andrade, A N . , LuQui, I.J. and Rafols, J.A. (1974). The primate globus pallidus: A Golgi and electron rnicroscopic study. Eur. J. Pharmacol., 1575-93. François, C., Yelnik, J. and Percheron, G. (1987). Golgi study of the primate substantia nigra. II. Spatial organization of denciritic arborizations in relation to the cytoarchitectonic boundarïes and to the striatonigral bundle . J. Cornp. Neurol., 265:473-493. Freund, T.F.,Powell, J,F. and Smith, A.D. (1984). Tyrosine hydroxylase-immunoreactive boutons in synaptic contacts wiîh identified striatonigrals neurons, with particular reference to dendritics spines. Neuroscience, 13:1 189- 1216. Gerfen, C.R. (1985). The neostriatal mosaic. 1. Cornpartmentai organization of projections from the striaturn to the substantia nigra in the rat J. comp. Neurol.., 207:282-303. Gerfen, C.R. ( 1992). The neostriatal mosaic: multiple levels of cornpartmental organization in the basal ganglia. Annu. Rev. Neurosci.., 15:285-320. Gerfen, CR. (1995). Dopamine receptor function in the basal ganglia. Clin. Neurophamacol, 18: 162-172 Gerfen, CR., Baimbridge, KG. and Thibault, J. (1987a). The neostriatal mosaic: III. Biochemical and developmental dissociation of patch-matrix mesostriatal systems. J. Neurosci, 7:3935-3944. Gerfen, CR., Herkenharn, M. and Miller, J.J. (1987b). The neostriatal mosaic: 11. Patch- and matrix- directed rnesostriatal doparninergic and non-dopaminergic systerns. J. Neurosci., 7:39 15-3934. Gibb, W.R. and Lees, A.J. (1991). Anatomy, pigmentation, ventral and dorsal subpopulations of the susbtantia nigra, and differentid ce11 death in Parkinson's disease. J. Neurol. Neurosurg. Psychiatry, 5:388-96. Grace, A.A. and Bunney, B.S. (1984a). The control of firing pattern in nigral doparninergic neurons: Single spike firing. Neurosci., 4:2866-2876. Grace, A.A. and Bunney, B.S. (1 984b). The control of firing pattern in nigrai doparninergic neurons: Burse firing. J. Neurosci., 4:2877-2890. Graybiet, A.M. (1978). Organization of the nigrotectal connection: An expenmentd tracer study in the cat. Brain Res., 143:339-348. Graybiel, A.M. ( 1990). Neurotransmitters and neuromodulators in the basal ganglia Trends Neurosci., 13 :244-253. Graybiel, A.M. and Ragsdale, C.W.Jr. ( 1978). Histochemistry distinct compartments in the striatum of human, monkey and cat demonstrated by acetycholinesterase staining. Proc. Natl. Acad. Sci. USA, 755723-5726. Gulley, R.L. and Wood, R.L. (1971). The fine structure of the neurons in the rat substantia nigra, Tissue and Cell., 3:675-690. HalIiday, G.M. and Tork, 1. (1986). Comparative anatorny of the ventromedid mesencephalic tegrnentum in the rat, cat, monkey and humain. J. Camp. Neural., 252:423-4-45, Hassani, O-K.,François, C., Yelnik, J. and Féger, J. (1997). Evidence for a doparninergic innervation of the subthdamic nucleus in the rat- Brain Res., 749:88-94. Hirsch, E.C. (1992). Why are nigral catecholaminergic neurons more vulnerable than other ceIIs in Parkinson's disease? Ann. Neurob , 32 Supp1:S 88-93. Hokfelt, T.,Everitt, B .J.. Theodorsson-Norheim, E. and Goldstein, M. ( l984a). Occurence of neurotensin-like immunoreactivity in subpopulations of hypothalamic, mesencephalic and medullary catecholarnine neurons. J. Cornp. Neural., 222543-560. Hokfelt, T., Lundberg, J.M., Schultzberg, M., Johansson, O., Skirboll, L., Anggard, A,, Fredholm, B., Harnberger, B., Pernow, B., Rehfeld, J. and Goldstein, M. (1980). CelIular localization of peptides in neural structures. Proc. R. Lond. B. BioL Sci., 2 10: 63-77. HoBeIt, T., Matensson, R,, BjorkIund, A., KIeniau, S. and Goldstein M. (I984b). Distribution maps of tyrosine hydroxyIase-immunoreactive neurons in the rat brain. In: A. Bjorklund and T.HokfeIt (eds.): Handbook of Chenrical Neuroanatomy, Vol. 2, Elsevier New York, pp.227-379. Horikawa, K. and Armstrong, W.E. (1988). A versatile means of intracellular Iabeling: Injection of biocytin and its detection with avidin conjugates. J. Neurosci Meth., 25: 1-11. Hsu, S.M., Raine, L, and Fanger, H. ( 1981). Use of avidin-biotin-peroxydase cornplex (ABC) in immunoperoxydase techniques: a cornparison between ABC and unlabelIed antibody (PAP) procedures. J. Hisrachem. Cytochern., 2 1 557-580- C.J. and Groves, PM. (1977). The substantia nigra of the rat: A Golgi Juraska, J.M., study. J. Comp. Neurol., 172:58S-6ûû. Kita, H., Kosaka, T,and Heizmann, C,W. (1990). Parvalbumin-immunoreactive neurons in the rat neostriatum: A light and electron microscopie snidy. Brain Res., 536: 1- 15. Lanca, A.J., Boyd, S., KoId, BE. and van der Kooy, D. (1986). The development of a patchy organization of the rat striatum. Brain Res., 392: 1- 10. Langston, J.W. (1985). Mechanisms of M P V toxicity: more answers more questions. Trends Pannacol. Sci, 375-378. Lavoie, B. and Parent, A. (1991). Dopaminergic neurons expressing calbindin in normal and parkinsonian monkeys. Neuroreport, 2:60 1-6O4. Lavoie, B.. Smith, Y.and Parent, A. ( 1989). Dopaminergic innervation of the basai ganglia in the squirrel monkey as revealed by tyrosine hydroxylase immunohistochemistry. J. Comp. Neurol., 288:36-52. Lévesque, M., Gagnon, S., Parent, A. and Deschênes, M. (1996). Axond arborization of corticostriatal and corticothalarnic fibers arising from the second somatosensory area in the rat. Cerebral Cortex, 6:759-770, Lindvail, 0. and Bjorklund, A. (1979). Dopaminergic innervation of the globus pailidus by collaterals from the nigrostriatai pathway. Brain Res., 172: 169-173. Meibach, R.C. and Katzman, R. (1979). Catecholaminergic innervation of the subthalamic nucleus: evidence for a rostral continuation of the A9 (substantia nigra) dopaminergic ce11 group. Brain. Res., 173:364-368. McCrodden, J.M., Tipton, K.F.,Sullivan, J.P. (1990). The neurotoxicity of MPTP and its relevarice to Parkinson's disease. Pharmacol. Toxicol., 67:8- 13. Mettler, EA. (1943). Extensive unilateral cerebral removals in the primate: Physiologie effects and resultant degeneration. J. Comp. Neurol., 79: 185-243. Nakanishi, H., Hori, N. and Kastuda, N. (1985). Neosuiatal evoked inhibition and effects of dopamine on globus pallidus neurons in rat slice preparations. Brain Res., 358:282-286. Nobin, A. and Bjorklund, A. (1973). Topography of the monoamine neuron systems in the human brain as revealed in fetuses- Acta Physiol. Scand., (suppl,) 388 :1-40. Oertel, W.H., Nitsch, C. and Mugnaini, E. (1984). Immunocytochemical demonstration of the GABA-ergic neurons in rat globus pailidus and nucleus entopedoncularis and their GABA-ergic innervation. In: R.G. Hassler and J.F. Christ (eds.). Parkinson-Specifc Motor and Mental Disorders (Adv. Neurol., Vol. 40), Raven Press, New York, pp.9 1-98, Oertel, WH., Tappaz, ML., Beord, A. and Mugnaini, E. (1982). Two-color irnmunohistochernistry for dopamine and GABA neurons in rat substantia nigra and zona incerta. Brain Res. Bull., 9:463-474. Olanow, C.W. (1987). Oxidation reactions in Parkinson's Disease. Neurology, 40:32-37. Parent, A. (1986). Comparative Neurobiolugy of the Basal Ganglia, Wiley, New York. Parent, A. (2996). Carpenter S Human Neuroanatomy, Wiliams and Wilkins, Baltimore. Parent, A., Bourassa, J. and Deschênes, M. (1996). The thalarnostnatal projection system: A single axon labeling study in rodents. In: C. Ohye, M. Kimura and J.S. McKenzie (eds). The Basal Ganglia V, Plenium Presss, New York, pp.3-10. Parent, A. and Cicchetti, E (1998). The current mode1 of basal ganglia organization under scrutiny. Mov. Disorders, 13: 199-202. Parent, A., Côté, P-Y.and Lavoie, B. (1995). Chernical anatomy of primates basal ganglia. Progress in NeurobioIogy, vo1.46: 13 1- 1%.' Parent, A. and Hazrati. L.-N. (1995). Functiond anatomy of the basal ganglia. 1. The corticobasai ganglia thdamo-cortical loop. Brain Res. Rev., 2O:g 1- 127. Parent, A., Lavoie, B., Smith, Y. and Bédard, P. ( 1 990). The dopaminergic nigropallidd projection in primates: distinct cellular ongin and relative sparing in MPTP-treated monkeys. In: M.B. Streifler, A.D. Korczyn, E. Melamed and Therapy. Advances in Neurology, Vol. 53, Raven Press, New-York, pp. 11 1-1 16. Parent, A., Mackey, A. and De Bellefeuille, L. (1983). The subcortical afferents to caudate nucleus and putarnen in primate: A fluorescence retrograde double labeling study. Neuroscience, 10:1137-1150. Parent, A. and Lavoie, B. (1993). Dopaminergic innervation of the basal ganglia in normd and parkinsonian monkeys. In: J. S. Schneider and M. Gupta (Eds.): Current Concept NI Parkinson 's Disease Research, Hogrefe and Huber, Toronto, 1993, pp. 403414. Paxinos, G . (1985). The Rat Nervous Systern (2nd Ed), Academic Press, San Diego. Paxinos, G. and Watson, C. (1986). The Rat Brain in StereotMic Coordinates, (2nd Ed), Academic Press SydneyPoirier, L.J., Giguère, M. and Marchand, R. (1983). Comparative morphology of the substantia nigra and ventral tegmental area in the monkey, cat and rat. Brain Res. Bull., 11:37 1-397. Rafols, I.A. and Fox, C.A. (1976). The neurons in the primate subthalamic nucleus: A Golgi and electron microscopic study. J. Comp. Neurol., l68:75- 112. Reiner, A. and Anderson, K.D. (1990). The patterns of neurotransmitter and neuropeptide CO-occurencearnong striatd projection neurons: Conclusions based on recent findings. Brain Res. Rev., 1S:2S 1-265. Resibois, A. and Rogers, J.H. (1992). Cdretinin in rat brain: an immunohistochemical study. Neuroscience, 46: 101-34. Rinvik, E., Grofova, L, Hammond, C., Féger, I. and Deniau, J.M. (1979). A study of the afferent connections to the subthalarnic nucleus in the monkey and the cat using the HRP technique. In: L.J. Poirier, T.L. Sourkes and P.J. Bédard (eds.). Advances in Neurology. Vol. 24, The Exrrapyramidal System and its Disorders. Raven Press, New York, pp.53-7 1. Rodrigo, J., Femandez P., Bentura, M.L., de Velasco, J.M.. Serrano, J., Uttenthal, 0. and Martinez-Murillo, R. (1998). Distribution of catecholaminergic afferent fibres in rat globus pallidus and their relations with cholinergie neurons. J. Chem. Neuroanat., 15: 1-20. Rogers, J.H. (1992). ImmunohistochemicaI markers in rat brain: Colocalization of cdretinin and calbindin-D28k with tyrosine hydroxylase. Brain. Res., 587:203-2 10 Schneider, J.S. and Dako, S. (199 1). Relative sparïng of the dopaminergic innervation of the globus pallidus in monkeys made hemi-parkinsonian by intracarotid MPTP infusion. Brain Res., 556:292-296. Smith, Y and Parent, A. (1986). Neuropeptide Y-immunoreactive neurons in striatum of cat and rnonkey: Morphological characteristics, inhinsic organization and CO-localization with somatostatin- Brariz Res,, 372:24 1-252- Smith, Y.and Parent, A. (1988). Neurons of the subthdamic nucleus display glutamate but not GABA irnrnunoreactivity. Brain Res., 453:353-356. ' and Descames, L. (1987). Distribution of GABA-immunoreactive neurons in the basal ganglia of the squirrel monkey (Sairnin sciureus). J. CompNeurol., 25950-64. Smith, Y., Parent, A., Séguéla, Smith, Y., Lavoie, B., Dumas, J. and Parent, A. (1989). Evidence for a distinct nigropallidal dopaminergic projection in the squirrel monkey. Brain Res., 48238 1-386. Szabo, J. (1980). Distribution of striatal afferents frorn the mesencephalon in the cat. Brain Res., 21:3-21. Ungerstedt, U. (1971). Stereotaxic mapping of the monoamine pathways in the rat brain. Acta Physiol. Scand. (suppl.), 367: 1-48. Wichmann, T. and DeLong, M.R. (1996). Functional and pathophysiological models of the basal ganglia C m Op. Neurobiol-, 6:75 1-758. Yarnada, T.,McGeer, PL., Baimbridge, KG. and McGeer, E.G. (1990). Relative sparing in Parkinson's disease of substantia nigra dopamine neurofis containing caibindin D-28K. Brain Res., 526:303-307. Yelnik, J., François, C., Percheron, G . and Heyner, S. (1987). Golgi study of the primate substantia nigra. 1. Quantitative morphology and topography of nigrai neurons. J. Comp. Neurol., 265:455-472.