chap2

Chap 2- L'immunité adaptative, prolongement de l'immunité innée
Lorsque la réponse innée ne suffit pas, des modifications apparaissent ds le sérum (cf poly Monsieur X) : par exemple,
AC et les lymphocytes sont plus nombreux. On sait que des CPA ( Cellules Présentatrices de l'Ag ayant phagocyté l'AG)
issues de la réponse innée entrent en contact avec des lymphocytes dans les organes lymphoïdes secondaires. Comment
sont fabriqués les anticorps ? Quel est leur rôle ? Quels sont les mécanismes mis en jeu avec les lymphocytes ?
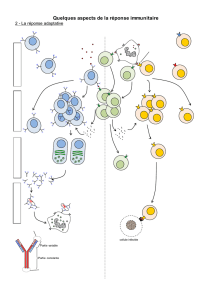
I- Les anticorps : des molécules solubles ds le milieu extracellulaire ou « humeurs »: ou réponse adaptative
humorale :
A) Détection des AC : cf TP1- Electrophorèse de protéines sériques et test d'agglutination- cf schémas TP1
4 méthodes principales : électrophorèse de protéines sériques, test d'agglutination, test Elisa et test d'Ouchterlony, à
comprendre en TP
B) Structure et fonction des AC :
Un anticorps ou gamma -globuline a une forme en Y. Sa
structure lui permet de fixer 2 antigènes .
2 chaines lourdes identiques entre-elles et 2 chaines légères
identiques entre-elles.
Chaînes légères et lourdes reliées par des ponts disulfures.
Une extrémité des bras courts du Y = Fab
= antigen binding fragment et contient une région variable.
Le reste des chaines est constant = fragment constant Fc.
Cet AC est libre, soluble, forme circulante dans le plasma.
C) Origine et rôles des AC : ex bac - les plasmocytes et lymphocytes B ex poly Mosier et p 303, 302
1. Les Ac proviennent de plasmocytes eux- mêmes provenant de la différenciation de certains LB. Ces LB
portaient exactement les mêmes AC mb que ceux secrétés dans le milieu intérieur par les plasmocytes (
ultrastructure adaptée à leur fonction, très riche en REG car fabrique les AC, protéines, en gde qté).
2. Chaque LB ne porte qu’un seul type d’AC et il existe au départ une grande diversité de LB ds le corps =
répertoire immunologique inné. Cette diversité existe avant tout contact avec l’antigène ( processus de
maturation des gènes particulier).
3. Seuls les LB dont les AC mb sont spécifiques de l'Ag sont sélectionnés par les AG directement et se
multiplient pour former un clone ( expansion clonale) . Certains LB deviendront mémoire tandis que les
autres se différencient en plasmocytes secréteurs d’AC anti-AG . Même si quelques LB spécifiques de l'AG
préexistent avant le premier contact, la sécrétion d’anticorps anti-Ag circulants ou libres est réalisée
qu’après l’entrée de l'AG ; il s’agit bien d’une réponse immunitaire adaptative + lente que la réponse
innée. Environ 8 jours pour cette différenciation ( = délai), et ganglions enflent.
4. L’expansion clonale et la différenciation nécessite la présence des LT et des macrophages ; on parle de
coopération cellulaire. ( médiateurs chimiques interleukines)
5. La liaison Ag- AC entraîne la formation de
complexes immuns. La formation de
complexe immun entraîne la neutralisation de
l’AG mais pas sa destruction. Savoir dessiner
un complexe immun. Cf Test d'agglutination
6. La destruction peut être assurée par
phagocytose facilitée ( savoir faire croquis).
Les phagocytes, macrophages et
polynucléaires, expriment des récepteurs de
la partie constante FC des AC, qui se fixent
sur les complexes immuns et les éliminent
par phagocytose dite facilitée ( opsonisation) .
7. Rem il existe aussi une destruction par action
lytique des protéines du complément qui sont
présentes dans le sérum à l'état soluble en
permananence.
Conclusion : On retient que les anticorps libres ( issus de LB/plasmocytes) surveillent le milieu
extracellulaire. C'est la réponse ou médiation humorale.

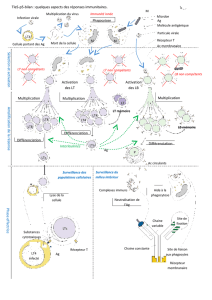
II- Les Ltc : une réponse adaptative cellulaire qui lutte contre les cellules infectées : ex p 304, 305, 306.
La destruction se réalise
par contact entre récepteur
T ( TCR) d'un Ltc,
lymphocyte cytotoxique
spécifique et la cellule
infectée – savoir le schéma
Les cellules infectées
présentent à leur surface
des fragments peptidiques
issus de protéines
virales associées au CMH
que ne présentent pas les
cellules saines ; elles sont
dès lors reconnues comme
cellules étrangères
( infection virale,
endobactéries, cancers...)
Les Ltc proviennent des LT CD8; Les LT8 par leurs récepteurs T spécifiques reconnaissent les cellules modifiées. La
production de LTC spécifiques à partir de LT précytotoxiques repose sur des étapes voisines de celles conduisant à la
synthèse de plasmocytes : sélection des LT 8 spécifiques de l’AG, multiplication clonale et différenciation). Les Ltc
portent des récepteurs T identiques à ceux des LT8 initialement sélectionnés par les CPA.
Ccl : Les Lt CD8 sont donc spécialisés par leur structure ds la surveillance des populations cellulaires. Les LTc
ont les mêmes récepteurs que les LT8 sélectionnés et sont produits au bout de quelques jours ( délai nécessaire).
Rem 1: La destruction in vitro des c infectées ne peut se réaliser que par des LT prélevés chez une souris ayant été
préalablement mise en contact avec le même virus, elle ne se produit pas si les LT st prélevés sur une souris n’ayant pas
rencontré l’agent pathogène donc les Ltc ne préexistent pas avant l'entrée d'un AG, ils sont donc les effecteurs également
de l’immunité adaptative.
Rem 2 : La grande diversité des lymphocytes est due à une production aléatoire de gènes remaniés ce qui permet de
répondre à une infinité d'AG potentiels alors qu'il n'y a pas une infinité de gènes. La maturation du système I résulte
d'un équilibre dynamique entre la production de cellules immunocompétentes et la répression des cellules
autoréactives. Si cet équilibre est déréglé, des maladies auto-immunes peuvent apparaître.
III- La coopération cellulaire entre les GB : ex p 308, 309, 310.
●La synthèse des effecteurs de la RIA , anticorps et Ltc, nécessite la présence des macrophages, et des
LT4.
●Les LT4 auxiliaires sont les seuls à fabriquer les interleukines Il2, molécules, peu spécifiques en elles-
mêmes, qui stimulent la multiplication des LB et des LT8 qui ont été en parallèle sélectionnés et activés (
= synthèse de récepteurs aux IL2) par le même antigène.
●Les LT4 auxiliaires ou helpers proviennent de certains LT4 spécifiques et sélectionnés par les CPA.
●A la suite de la phagocytose d’un antigène dans l’organisme, des cellules dendritiques ou des
macrophages sont devenues des CPA ( cellules présentatrices de l'AG porteuses d'un CMH modifié avec
des fragments antigéniques que n'ont pas les cellules saines) et migrent ds ggl lymphatiques. Les CPA ou
les cellules infectées sont alors reconnues par certains clones naïfs de LT4. Certains se différencient en
LT4 helpers, d'autres en LT4 mémoires.
Conclusion : les interleukines Il2 permettent l'amplification clonale des LB et des LT, puis la différenciation en
effecteurs : AC anti -AG libres et LTc spécifiques de l'AG. Comme les LT4 sont les seuls à fabriquer les IL2, les
Lt4 sont considérés comme les chefs d’orchestre de la réponse immunitaire acquise. Ds le cas du SIDA, le
virus infecte des cellules cibles qui sont les LT4 et les macrophages qui sont détruits progressivement. La chute
du tx des LT4 de toutes spécificités en phase terminale sous un seuil de 200/ mm3 de sang entraine
l'insuffisance d'IL et donc de l'amplification et de la différenciation des effecteurs de l'immunité adaptative. Donc
apparition de maladies opportunistes. Trithérapies possibles. Connaître les phases du SIDA cf graphes p 308.
Alors que l'i innée est largement répandue chez les EV ( 800 MA) , I adaptative est propre aux vertébrés
( 400 MA). Elle s'ajoute à l'I innée dont elle est un prolongement et assure une réponse plus lente mais plus
spécifique contre l'AG. Elle permet la synthèse d'effecteurs : Ac libres et LTC qui ne préexistent pas avant
le 1er contact avec Ag. Lors de la phase de destruction de l'AG, les 2 immunités sont associées. I innée
intervient donc au début et à la suite de l'I adaptative.
1
/
2
100%