30/10/13 LIU Alice L3 NGUYEN Aude L3 Système

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Physiologie de la vision
30/10/13
LIU Alice L3
NGUYEN Aude L3
Système neurosensoriel et psychiatrie
Dr Trebuchon-Da Fonseca
20 pages
Physiologie de la vision
INTRODUCTION : Rappels sur les signaux visuels et la lumière :
Les stimulis visuels sont des ondes électromagnétiques avec une longueur d’onde particulière : le spectre du
visible va de 400 à 700 nm. L'Homme est capable de distinguer environ 200 nuances de couleurs avec un
seuil de différence de 1,5 nm.
Cependant, la sensibilité du système visuel dépend aussi de l’éclairement. Dans l'obscurité il y a un tas de
choses que l'on ne perçoit plus, notamment la couleur.
On parle de vision :
–chromatique (à fort éclairement) où la lumière est
codée.
–achromatique ou scotopique (à faible éclairement)
où seule la distinction noir / blanc est faite.
Si on regarde cette image, à partir de taches noires et
blanches, on voit une information se dessiner : un dalmatien.
La perception dépend de ce que l’on sait, de ce que l'on a en
mémoire... On perçoit le monde à travers des lunettes qui
nous sont propres (apprentissage, images, ce qu’on peut
connaître). → C'est l' effet topdown (utilisation de
l’information que l’on connaît pour décoder l’information
perceptive). Si on n’a jamais vu de chien, on ne peut pas voir le dalmatien sur cette image.
1/20
Plan
A. L'oeil
I. Rappels anatomiques de l'oeil
II. Formation de l'image sur la rétine
III.La rétine
B. Transfert à travers les voies optiques centrales (voie rétino-géniculo-corticale)
I. Anatomie
II. Organisation des voies optiques, notion de champ visuel
III. Corps genouillé latéral du thalamus
C. Traitement de l'information corticale
I. Cortex visuel primaire
II. Cortex visuel associatif
D. Explorations neurophysiologiques : ERG et PEV
I. ERG flash
II. Potentiel évoqué visuel
III. Potentiel évoqué cognitif

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Physiologie de la vision
A. L'oeil
L'oeil permet de transformer la lumière (onde électromagnétique) en message nerveux (potentiel d'action, PA).
I. Rappels anatomiques de l'oeil
L'oeil est constitué de plusieurs segments :
–la chambre antérieure, entre la cornée et le cristallin
–la chambre postérieure avec l'épithélium pigmentaire à la face interne, puis la rétine sur l’épithélium
pigmentaire. Les informations partent ensuite au niveau du nerf optique.
II. Formation de l'image sur la rétine
L’œil reçoit des rayons lumineux émis ou réfléchis par des objets de l'environnement. Le premier traitement des
informations visuelles permet de focaliser l'image sur la rétine dans un endroit particulier : la fovéa.
Pour que l'image se forme sur la rétine, il y a traitement de l’information par des lentilles convergentes, cornée
et cristallin, qui font converger les signaux lumineux sur des points précis de la rétine.
1. Réfraction par la cornée :
La cornée est une lentille convergente qui va permettre une réfraction des signaux lumineux de 42 dioptries. Si
la convexité cornéenne est anormale, l’image se forme soit trop tôt (avant la rétine) : myopie, soit après la
rétine : hypermétropie.
2. Adaptation à la luminosité grâce au réflexe pupillaire de l'iris
L’iris est situé entre la cornée et le cristallin. C'est un muscle lisse qui permet le contrôle du diamètre de la
pupille. Il y a 2 types de muscles :
•muscles circulaires qui permettent à l'iris de se contracter : myosis (environnement lumineux)
•muscles radiaires qui permettent à l'iris de s'écarter : mydriase (obscurité)
2/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Physiologie de la vision
Ex : la dilatation pupillaire en ophtalmologie est très inconfortable => éblouissement permanent car il n'y a
plus de contraction de l'iris.
Il n'y a pas de contrôle conscient des muscles de l'iris. C'est le système nerveux végétatif qui s'en charge : le
système orthosympathique assure la mydriase alors que le parasympathique permet le myosis.
3. Accommodation par le cristallin :
Le cristallin est suspendu par les ligaments suspenseurs des 2 côtés. Il adapte le signal notamment pour la
vision de près. C’est un système dynamique (à l'inverse de la cornée) qui permet l’accommodation. Il ajoute
12 dioptries aux dioptries précédentes de la cornée.
Ex : presbytie = absence d’accommodation du cristallin. Elle est compensée par des lunettes.
III. La rétine
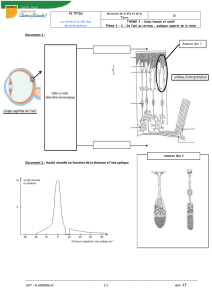
a. Anatomie de la rétine
Elle possède une organisation laminaire en 3 couches :
–Partie distale : couche de photorécepteurs (cônes et bâtonnets)
–Partie moyenne : cellules bipolaires + cellules horizontales peu visibles
–Partie proximale (+ proche de la lumière) : cellules ganglionnaires et départ des axones vers le nerf
optique
À la partie la plus externe de la rétine se trouve l’épithélium pigmentaire qui joue un rôle dans la reconstruction
de certaines parties des photorécepteurs.
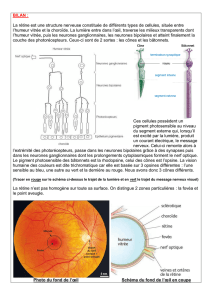
La distribution des récepteurs au sein de la rétine n’est pas la même partout.
–Il n'y a pas de bâtonnets au niveau de la fovéa, ils se répartissent tout autour.
– Il y a cependant un nombre très important de cônes dans la fovéa, c’est la zone d’acuité maximale
de l’œil.
La fovéa se trouve dans une partie un peu plus étendue appelée macula.
3/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Physiologie de la vision
b. Les photorécepteurs :
Il y a 2 types de photorécepteurs : les cônes et les bâtonnets.
Ces 2 types de cellules diffèrent dans la partie terminale de leur
structure mais elles sont constituées à peu près de la même façon :
–Segment externe comportant un empilement de disques dans
lesquels sont placés les photopigments
–Segment interne avec un corps cellulaire et une terminaison
synaptique (synapse avec les cellules bipolaires)
Il y a 120 millions de bâtonnets et 6 millions de cônes dans chaque œil, c'est-à-dire 20 fois plus de bâtonnets
que de cônes dans la rétine humaine. Les cônes sont très importants au niveau de la fovéa et les bâtonnets au
niveau de la rétine périphérique.
Les muscles oculomoteurs et la mobilité rapide de la tête permettent de toujours fixer l'image sur la fovéa qui
permet d'avoir l'acuité la plus fine possible grâce aux cônes.
Les bâtonnets sont 1000 fois plus sensibles à la lumière que les cônes. Ils sont responsables de la vision
scotopique (de nuit).
Les cônes sont responsables de la vision photopique (jour). Ils permettent le codage de la couleur. Il y a trois
grands types de cônes avec des pigments différents, spécialisés dans la vision des couleurs (bleu, vert et rouge).
Les photopigments sont sensibles à une longueur d’onde particulière.
4/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Physiologie de la vision
c. Phototransduction +++
Phototransduction = l’énergie lumineuse électromagnétique est transformée en potentiel de membrane.
(Attention : il n'y a pas de PA à cette étape là !)
La transformation est possible grâce au photopigment, qui est l'association de deux molécules : opsine et
rétinal (ou rétinène).
Le rétinal existe sous deux formes stéréochimiques selon une interaction particulière qui se fait grâce à la
lumière : la forme cis inactive et la forme trans active.
Opsine + rétinène = rhodopsine.
La rhodopsine permet au photorécepteur de traiter l'information.
1) Le photon lumineux arrive sur la rhodopsine. Le rétinène passe alors de la forme cis à la forme trans. Il y a
donc une modification des sites de liaison de la rhodopsine qui active elle-même une protéine G : la
transducine.
2)La transducine stimule une phosphodiestérase qui hydrolyse le GMPc en GMP. La concentration de GMPc
au sein de la cellule diminue.
3) A la surface du photorécepteur, il y a des canaux sodiques GMPc-dépendants. Lorsque le taux de GMPc
diminue, les canaux Na+ se ferment → entraînant une hyperpolarisation.
La nuit
Dans le segment externe du photorécepteur, les canaux Na+ sont ouverts et il y a entrée constante de Na+ grâce
à une forte concentration de GMPc. Ceci entraine une dépolarisation partielle permettant à la cellule de garder
un potentiel stable aux alentours de -40mV. Dans le segment interne il y a une fuite de potassium vers l'espace
extracellulaire.
5/20
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%