plasticite synaptique NAASSILA 021104 poly

1
Plasticité neuronale
Martine Daoust, PU
Olivier Pierrefiche, MCU
Mickaël Naassila, MCU, [email protected]
http://www.u-picardie.fr/sante/pagesliees/grap/accueilgrap.html
JE 2462
ETUDE DES BASES NEUROBIOLOGIQUES
DE L’ALCOOLODEPENDANCE
I. Animal:
Syndrome d’alcoolisme fœtal
Exposition précoce => vulnérabilité ultérieure
Approches:
cellulaire et moléculaire
électrophysiologique
comportementale
Souris KO, sélection génétique
II. Clinique:
Pharmacogénomique (étude d’association
polymorphismes génétiques (neurotransmission) &
réponse aux traitements)
Plasticité neuronale
Plasticité neuronale
Plasticité fonctionnelle & morphologique
Capacité à adapter son activité (et/ou l’architecture
des réseaux) en fonction du niveau de stimulation…
Plasticité neuronale
Plasticité synaptique
Efficacité de transmission synaptique
Force de transmission synaptique
Solidité des connexions synaptiques
PROLIFERATION
DIFFERENCIATION
MIGRATION
SYNAPTOGENESE
REMODELAGE (nombre neurones/connexions)
Plasticité neuronale
Établissement de la polarité neuronale in vitro. Dans les neurones en
culture, immédiatement après l’attachement au substrat, on observe la formation
de lamellipodes puis l’apparition des prolongements non différenciés, les
neurites. L’un de ces neurites, le futur axone, va croître plus rapidement: le
neurone acquiert sa polarité. Le développement des dendrites et de l’axone se
poursuit alors jusqu’à la maturité.

2
Cônes de croissance
‘têtes chercheuses’ Remodelage fonctionnel
Seules les synapses “actives” persitent,
rétraction
rétraction
perforation
Epine dendritique
Cou Tête
Spine
apparatus
Retic endo lisse
+ ribosomes
Densité
postsynaptique
1microm
Différentes morphologies des épines
mushroom
shaft
concave
sessile
Thiny
1microm
Reconstructions tri-dimensionnelles
Epine branchée épine à synapses multiples
fenestrée Fer à cheval segmentée
Segment dendritique
Boutons, varicosités…
PSD en
rouge
1microm 1microm
Role des épines et de leur diversité
1- microcompartiment relativement isolé de la dendrite
(conserve l’individualité des ‘inputs’) peut donc fonctionner
comme une unité intégrative multifonctionnelle
2- contient des éléments régulateurs pour effectuer des
processus biochimiques spécifiques
3- évite une propagation non spécifique de la potentialisation
(spécificité de la LTP)
4- structure & fonction modulées selon l’activité synaptique
locale
5- constriction du cou peut engendrer une barrière résistive…
6- nombre de vésicules, surface de libération
7- perforation augmente le nombre de sites de libération
Plasticité neuronale Plasticité neuronale (1)
Retard mental……………………….
Retard mental associé avec des
altérations dendritiques et
épines dendritiques.
Réduction du nombre et épines
longues et fines (immatures)
avec épines ‘champignons’
Dysgenésie des épines.
6 mois 12 ans

3
Acides aminés neurotransmetteurs
Na+= 14
K+ = 160
Ca2+ = 10-4
Cl-= 14
Cl-Na+, Ca2+ (sortie K+)
Na+= 140
K+ = 3-4
Ca2+ = 1
Cl-= 150
mmol/l
Hyperpolarisation Dépolarisation
•GABA
•glycine
•β-alanine
•taurine
•glutamate
•aspartate
3Na+
2K+
ATPase
•Canaux
•Récepteurs canaux
•+(métabotropiques)
Le neurone: l’unité
fonctionnelle de la mémoire
Plusieurs centaines de milliards de
neurones
Chacun relié à dix ou vingt mille autres par
des connexions appelées synapses
Plus d’un million de milliards de
connexions
Plasticité neuronale
Plasticité neuronale
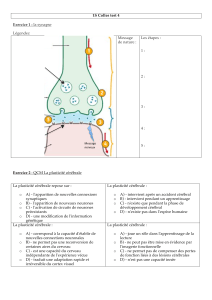
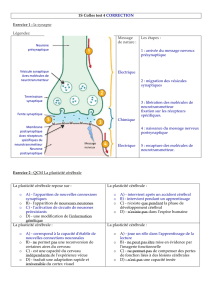
Synapse = point de « connexion » entre deux neurones
1 mm3de substance grise du cortex peut contenir 5 milliards de synapses.
chimique
électrique neuromusculaire
Permet transfert unidirectionnel
de l’information entre neurones
La ‘big
picture’
INACTIVATION
Dégradation
Capture
autorécepteurs

4
Plasticité neuronale
Représentation
schématique des
différentes
modalités
d’interaction entre
les neurones et le
réseau astrocytaire.
Neurotransmission chimique
La nature de la réponse post-synaptique dépend:
Type de récepteur
Inhibitrice ou excitatrice
L’amplitude de la réponse au NT qui elle-même dépend de:
•quantité de NT libéré
•nombre de récepteurs
•état des récepteurs
‘Simpliste’ ‘réaliste’
100 milliards neurones
~1 million de milliards
connexions (synapses)
Plasticité neuronale
Environ 3000 boutons par neurone !
PPSE PPSI
Potentiel d’action
INTEGRATION NEURONALE
Sommation spatiale &
temporelle des signaux
probabilité
Plasticité neuronale
Densité post-synaptique

5
Plasticité neuronale
Densité post-
synaptique
2 types de récepteurs des NT
IONOTROPIQUES :
Liaison NT induit un changement conformation du récepteur qui
entraîne l’ouverture d’un canal ionique intrinsèque, permettant aux
ions de circuler en descendant leur gradient électrochimique
Influx de cations (Na+, Ca2+)ÎDEPOLARISATION (PPSE)
Influx de Cl-(ou efflux de K+) ÎHYPERPOLARISATION (PPSI)
METABOTROPIQUES :
Liaison NT induit un changement conformation du récepteur qui entraîne la
liaison d’une protéine G et la production de seconds messagers (AMPc, GMPc,
IP3, DAG, Ca2+ => conductance ionique, activation de protéine kinase, libération
de Ca2+ des stocks intracellulaires ou liaison à des facteurs de transcription) après
activation de systèmes enzymatiques.
La réponse est lente à initier et de longue durée comparativement aux récepteurs
ionotropiques
2 types de récepteurs des NT
Récepteurs métabotropiques & ionotropiques sont des protéines
qui n’ont pas d ’homologie structurale
Caractéristiques générales des récepteurs ionotropiques
•membres de familles de protéines ~ tx d’homologie
•complexes protéiques avec des régions spécifiques
traversant la mb
•activation très rapide (~sub-milliseconde)
•sélectivité ionique (eg. cation vs. anion)
•la +part désensibilisation rapide après exposition à 1
agoniste
Principales familles de récepteurs ionotropiques
•nAChR, GABAA, glycine, sérotonine-3 (5-HT3)
•récepteurs Glutamate (NMDA, kaïnate, AMPA)
ROLE CLE DE CES RECEPTEURS DANS LA
PLASTICTE SYNAPTIQUE…
Ancrage historique
1894: Santiago Ramòn y Cajal propose l’existence de la genèse
neuronale de synapses. Théorie de la mémorisation : “c’est la
croissance de nouvelles connexions qui permet le stockage”
1949: Hebb émet l’hypothèse que les connexions synaptiques
peuvent augmenter leur efficacité de transmission électrique.
“les neurones qui déchargent en même temps établissent des
connexions privilegiées”
1970: Kandel confirment l’hypothèse de Hebb en étudiant
l’aplysie (limace).
1973: Bliss et Lomo démontrent la plasticité neuronale.
Plasticité neuronale
apprentissage
mémoire
Acquisition et mise en réserve de
l’information par suite de
l’expérience
Forme relativement permanente de
la mise en réserve de l’information
apprise
Dépend: attention, répétition, motivation,
organisation
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
1
/
27
100%