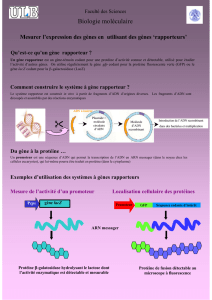
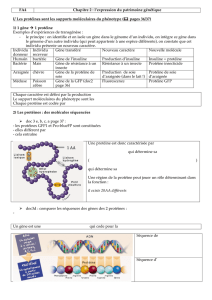
Génie génétique, production de médicaments et - iPubli

2

�
t
t
® \..
.\
@
, I
�
.
'
86

BRÈVES
1
/
3
100%