MASMOUDI Sounda et POREE Solène 11 octobre 2010 Biochimie

MASMOUDI Sounda et POREE Solène
11 octobre 2010
Biochimie digestive, Exploration biochimique du pancréas endocrine, Véronique David
Diaporama disponible sur le réseau
EXPLORATION BIOCHIMIQUE DU
PANCREAS ENDOCRINE
I. Introduction
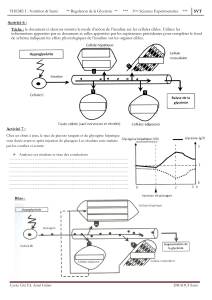
Le pancréas comporte deux parties :
une portion acineuse, ayant une fonction exocrine, qui sécrète des enzymes et des ions, dans
la lumière du tube digestif, pendant la digestion.
→ cette partie représente 98% de la glande
une portion glandulaire,ayant une fonction endocrine, située surtout au niveau de la queue
du pancréas.
→ cette partie représente 2% de la glande
Elle est constituée des ilots de Langerhans, comportant plusieurs types cellulaires :
▪ les cellules α ou cellules A (15 à 20% des cellules) situées à la périphérie, qui vont
sécréter le glucagon
▪ les cellules β ou cellules B (60 à 70% des cellules) situées au centre, qui vont
sécréter l'insuline
▪ les cellules δ ou cellules D (5 à 10% des cellules) qui vont sécréter la somatostatine
▪ les cellules F qui vont sécréter le polypeptide pancréatique
Le sang va circuler dans les ilots du centre vers la périphérie via un réseau de capillaires, puis va
dans la veine porte hépatique, ce qui va délivrer au foie la totalité des hormones pancréatiques
produites.
II. Insuline : hormone clé du métabolisme
organique
Le pancréas a une fonction essentielle : assurer l'homéostasie du glucose (donc le contrôle
de la glycémie).
C'est la résultante de plusieurs paramètres : l'absorption du glucose, production par le foie et
utilisation par les cellules organiques.
Ainsi le maintien de la glycémie se fait par un contrôle hormonal de la production hépatique
du glucose, grâce à deux enzymes pancréatiques : insuline et glucagon. Ces dernières ont un rôle
majeur → la pancréatectomie totale est incompatible à la vie.

A. Nature et biosynthèse
L'insuline est une hormone peptidique de 51 acides aminés (Aa) formée de deux chaines :
une chaine α, de 21Aa
une chaine β, de 30Aa
reliées entre elles par des ponts dissulfures
L'insuline peut exister sous plusieurs formes :
monomérique, biologiquement active en cas de concentration faible
di- ou héxamérique, en cas de concentration plus élevée
La synthèse de l'insuline est assurée dans les cellules β des ilots de Langerhans, en plusieurs
étapes :
1. le produit initial de la traduction des ARNm est la préproinsuline. Cette dernière a une
séquence signal de 25Aa, lui permettant une pénétration rapide au cours de la synthèse dans
le REG. Cette sequence sera ensuite clivée par une peptidase specifique.
2. On aura alors la proinsuline, qui quitte le REG par des microvésicules pour rejoindre le
GOLGI. A ce niveau, elle sera dirigée vers des vésicules ( recouvertes de clathrine ). Ces
vésicules sont de type contrôlé, car libérées seulement en réponse a un signal.
3. Les vésicules subiront ensuite une acidification de leur contenu. Seront alors libérées
insuline et peptide C, par exocytose, et en fonction des besoins.
Cette secretion d'insuline est biphasique :
une phase brève ( liberation d'insuline stockée en cas de besoins rapides )
une phase lente ( resynthese d'insuline en cas de prolongation de la stimulation )
L'insuline n'est jamais sécrétée seule, mais toujours avec le peptide C ( important pour les dosages
car il y a liberation d'autant de peptide C que d'insuline )
B. Action de l'insuline
L'insuline a deux actions : - métabolique ( agissant sur les glucides, lipides et proteines )
-de croissance
1. sur les glucides :
Action hypoglycemiante : rôle sur le transport du glucose
→ le glucose va dans les cellules via des transporteurs. Dans le cerveau, le foie, les globules rouges
et globules blancs ( consommateurs permanents de glucose ) les transporteurs ne sont pas controlés
par l'insuline. En revanche, les autres tissus ( en particulier les muscles et le tissu adipeux )
dépendent de l'insuline pour capter et utiliser le glucose.
L'augmentation plasmatique du glucose en cas de carence en insuline ( diabète ) empêchera
l'entrée du glucose dans les cellules périphériques. L'insuline est donc la seule hormone permettant
la pénétration du glucose dans les cellules.
La moitié du glucose est transformée en energie, l'autre moitié est stockée sous forme de
graisse.

L'insuline :
Stimule la glycogenogenese, en activant la glycogene synthetase
Inhibe la glycogénolyse
Stimule la glycolyse
Inhibition de la néoglucogenese
Stimule la synthèses proteique ( ce qui diminue la quantité de substrat disponible pour la
néoglucogénèse hepatique )
2. sur les lipides :
Action lipogenique :
→ active la lipoproteine lipase qui existe sur la paroi des capillaires, et qui permet l'hydrolyse des
triglycerides ( TG ), des VLDL et des chylomicrons. Cela entraine une liberation des acides gras (
Ag ) libres, captés par le tissu adipeux, qui va ensuite resyntheser des TG.
L'insuline :
Active les enzymes de la glycolyse : production de pyruvate et de glycerol necessaires a la
synthsese de TG
Inhibe la lipase hormono sensible et s'oppose alors a la lipolyse intra adipocytaire.
En cas d'absence d'insuline, les Ag libres sont convertis en corps cetoniques par le foies, ce
qui entraine l'acidocetose observée chez les diabetiques.
3. sur les protides :
Stimule l'entrée active des Aa et synthese des proteines dans la muscle et inhibe leur degradation.
Les effets seront differents selon les tissus :
si la concentration en insuline du plasma augmente, alors :
on aura stimulation de GLUT 4 au niveau du muscle, ce qui augmente la captation et
l'utilisation de glucose. Il y aura alors synthese de glycogene.
Stimulation de GLUT 4 au niveau des adipocytes, ce qui augmente la capacité de
captation du glucose et stimule la synthese en TG.
Augmentation de la captation de glucose par diffusion rapide dans le foie, ce qui stimule
la synthese de glycogene, de TG, en l'absence de formation de corps cetoniques.
On aura les effets inverses si la concentration en insuline du plasma diminue.
C. Mécanismes d'action

Pour être active, l'insuline se lie sur des récepteurs membranaires glycoprotéiques, existant
sur tous les types de cellules, mais leur nombre varie selon le tissus.
Structure des ces récepteurs : appartiennent à la famille des récepteurs à activité tyrosine kinase.
Ces récepteurs sont caractéristiques des facteurs de croissance IGF1 (qui a une grande homologie
avec le récepteur de l'insuline), EGF, PDGF.
Ce récepteur est un hétérodimère de deux sous unités α extra-cellulaires reliées à deux sous
unités β transmembranaires.
Chacune de ses sous unités à un rôle particulier :
α : reconnaissance de l'insuline, qui s'y fixe
β : protéine transmembranaire qui a activité tyrosine kinase, qui a site
d'autophosphorylation sur 3 tyrosines.
L'insuline transmet la signalisation grâce à plusieurs cascades de signalisation qui
interviennent sur des substrats spécifiques : substrat du récepteur à l'insuline (IRS).
→ phosphorylation de tyrosines kinases qui par phosphorylation vont activer les IRS
Effets cellulaires de l'insuline sont de différents types :
Quand l'insuline se fixe, on a autophosphorylation qui entraîne la stimulation de l'entrée du
glucose, par translocation des récepteurs transmembrainaires du glucose.
Stimulation de l'expression de gènes d'enzymes qui agissent sur ce métabolisme (rapidement
ou lentement)
D. Régulation de la sécrétion
Dans les conditions basales, la sécrétion d'insuline est de 0,2 à 1,5 UI/h.
À jeun, l'activité de l'insuline est très basse (2 à 17 mUI/L)
À l'inverse on observe un pic après le repas, suite à une stimulation par la présence de
glucose (10 à 100 mUI/L)
Réponse biphasique à la présence de glucose :
rapide (libération de l'insuline préformée)
lente (néosynthèse d'insuline si la stimulation perdure)
→ la concentration de glucose stimule donc la libération et la synthèse de l'insuline.
En dessous de 4,4 à 5,5 mmoles/L, il n'y a pas de sécrétion d'insuline (c'est la concentration
seuil de sécrétion).
Glucose pénètre dans les cellules, puis la transduction du signal glucidique semble se faire
par l'intermediaire du metabolisme glycolitique :
la glycolyse libere de l'ATP
cette augmentation d'ATP induit la depolarisation membrannaire, avec fermeture de
canaux K+ dépendant
Puis repolarisation membrannaire, permettant l'ouverture des canaux Ca2+
cette augmentation de Ca2+ va, par l'intermediaire de la proteine kinase C, libérer
l'insuline par exocytose.
La sécrétion de l'insuline n'est pas régulée que par le glucose, même si elle reste la
stimulation majeure.
En effet, on aura différentes stimulations :

Augmentation de la glycémie
Augmentation de la production d'Aa
Quantité de glucagon en exces
hormones gastro intestinales
augmentation de l'activité parasympatique ( PΣ )
A l'inverse, la diminution de la secretion d'insuline est entrainée par:
la diminution de la glycemie
la somatostatine
Adrenaline plasmatique
l'augmentation de l'activité orthosympatique ( OΣ )
contrôle de la biosynthèse : effectuée majoritairement par le glucose
En effet, le glucose :
stimule la traduction des ARNm de preproinsuline
stimule la traduction de genes
stabilise les ARNm
→ tout contribue à produire de l'insuline
Il existe aussi un contrôle au niveau des cellules cibles, car on y trouve des récepteurs à
l'insuline. Au niveau des cellules il existe un turnover important : 10000/h (donc on a un recyclage
permanent).
Après fixation de l'hormone, on a internalisation, puis l'insuline se défixe du récepteur qui sera
ensuite recyclé à la surface.
La production d'insuline est donc régulée très finement, et essentiellement par le glucose.
E. Dosage de l'insuline
Le dosage présente un intérêt en cas de :
diabète insulinodépendant (de type I) (DID) : dû à la destruction des cellules β des ilots
de Langerhans ; on aura donc plus de production d'insuline
diabète non insulinodépendant (de type II) (DNID) : les cellules deviennent résistantes à
l'insuline
Le dosage de l'insuline n'est pas intéressant pour le dépistage ou le diagnostic immédiat,
cependant ; il est important pour l'exploration de la capacité insulinosécrétoire du pancréas et le
dépistage de l'hypoglycémie.
1. Le dosage de l'insuline est perturbé par plusieurs éléments, comme par exemple une faible
quantité proinsuline non clivée et une quantité équimolaire de peptide C dans le sérum.
Le métabolisme et l'extraction de l'insuline et du peptide C sont différents, car le foie capte
50% de l'insuline plasmatique à chaque passage donc la demi-vie de l'insuline est de 4 minutes alors
que la demi-vie du peptide C est plus longue (30min) et donc reste plus longtemps dans le sang.
Peptide C est éliminé par le rein, donc son dosage est intéressant car plus stable.
2. Prélevement : le matin à jeun sur sérum non hémolysé (car hémolyse des GR
entraîne la libération d'enzyme qui dégrade l'insuline ce qui fausserait le dosage)
3. Interférences : dans les dosages il existe des interférences :
 6
6
1
/
6
100%