Réponse cytotoxique C`est le troisième mécanisme effecteur de la

Réponse cytotoxique
C'est le troisième mécanisme effecteur de la réponse immunitaire.
La réponse cytotoxique joue un rôle important dans la lutte anti-virale et anti-tumorale.
I) Reconnaissance
A) Les antigènes reconnus
Les antigènes reconnus sont les antigènes viraux, tumoraux (les tumeurs expriment des antigènes
qui sont des antigènes du soi modifiés) et des allo-antigènes (antigènes du non soi, portés par les
greffes).
B) Les mécanismes de reconnaissance
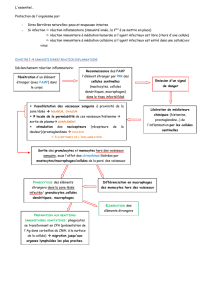
Les LTCD8 circulent sous une forme appelée pré-cytotoxique (préCTL). Ils vont avoir besoin d'une
série de signaux pour devenir cytotoxiques. Ils vont s'accrocher à la cellule cible par le système
LFA-1/ICAM-1, ils vont reconnaître l'antigène présenté par des molécules de classe I du CMH par
leur TCR. Le co-signal CD28/B7 est également nécessaire.
La plupart des cellules n'expriment jamais la molécule B7. Les cellules jouent un rôle important.
• Les LTCD8 peuvent être directement activés par les cellules dendritiques qui portent le B7, qui
peuvent être infectées. Si elles sont infectées par un virus ou un bactérie, l'antigène va être présent
dans le cytosol. Dans ce cas là, il y aura production d'IL-2 par la cellule dendritique, ce qui entraîne
une prolifération de la cellule dendritique et une différenciation en LTCD8 cytotoxique. A ce
moment là, le LTCD8 cytotoxique peut reconnaître d'autres antigènes, mais il ne nécessite plus de
cosignal.
• La cellule dendritique n'est pas infectée. Elle va utiliser le crosspriming ou cross présentation: la
cellule dendritique va internaliser des antigènes extracellulaires. On a découvert un système qui
permet à la cellule dendritique de faire exprimer un antigène extracellulaire, présent dans les
phagosomes, par les molécules de classe I du CMH. La membrane du phagosome comporte des
parties de membrane du réticulum endoplasmique, où on a des molécules de classe I et des
molécules du système TAP transporter. Des fragments vont sortir dans le cytosol et vont être
dégradés par le protéasome, réinsérés par le système TAP dans le phagosome, puis association avec
les molécules de classe I du CMH.
La cellule dendritique ou le macrophage peut rentrer en apoptose, ce qui forme des bulles
apoptotiques contenant des antigènes extracellulaires (→ crosspriming).
Si la cellule dendritique n'exprime pas suffisamment B7 et donc n'est pas capable d'activer le
lymphocyte T cytotoxique. Dans ce cas là, elle va avoir besoin des LTh1. La cellule dendritique va
exprimer l'antigène par les molécules de classe I et II du CMH. Le LTh1 va reconnaître l'antigène et
va augmenter l'expression de B7 sur la cellule dendritique. Au moment où elle exprime
suffisamment de B7, le LTCD8 passe du stade pré-cytotoxique au stade cytotoxique.
Les LTCD8 ont un énorme potentiel de cytotoxicité.
II) Comment le LTCD8 cytotoxique va tuer ses cellules cibles
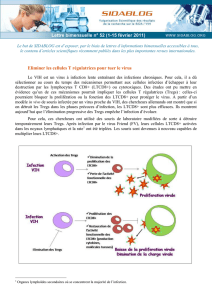
La cellule cytotoxique va adhérer à la cellule cible (cette adhésion nécessite la présence de
magnésium). Elle va procéder à un réarrangement: les organites et le noyau vont partir à l'autre bout

de la cellule (dans la région opposée à la zone de contact) et on voit apparaître des granules dans la
zone de contact. L'activation a conduit à la production des granules. C'est en parti le contenu des
granules qui va permettre la destruction de la cellule cible. Les granules vont déverser leur contenu
dans l'espace, puis le LTCD8 se détache, et la cellule cible meurt par apoptose (15 minutes à 3
heures).
Dans cette étape de destruction, il va y avoir plusieurs facteurs importants:
- les granules contiennent un certain nombre de substances, dont:
∙ la perforine: cytolysine qui se polymérise pour former un pore de 5 à 20 nm de
diamètre dans la membrane, et elle forme un complexe avec la serglycine et les
granzymes. Les granzymes vont pénétrer dans la cellule cible grâce à la perforine.
Les granzymes sont des sérine-estérases qui vont activer la caspase-3, qui active une
DNase (CAD), ce qui entraîne une fragmentation de l'ADN. Les granzymes activent
aussi BID, ce qui entraîne une fragmentation de la membrane des mitochondries,
avec une libération de cytochrome c (molécule pro-apoptotique).
∙ la granulysine
Les cellules apoptotiques sont éliminées essentiellement par les macrophages (→ pas d'activation
des macrophages, donc pas de réaction inflammatoire).
- le système Fas/FasL: la molécule Fas est exprimée par la cellule cible. Le LTCD8 activé va
exprimer FasL, et l'interaction avec Fas active des caspases et induit l'apoptose.
- cytokines (TNF-α et interféron γ): peuvent avoir un effet cytotoxique.
Quel que soit le mécanisme de lyse, les LTCD8 résistent car le passage des granules est ciblé vers la
cellule cible.
La reconnaissance d'une cellule cible infectée ou tumorale peut se faire de différentes manières:
- macrophage par système ADCC
- cellule NK par le missing-self
Une fois que la reconnaissance est faite, les mécanismes de destruction sont les mêmes pour la
cellule NK, le macrophage ou pour le LTCD8.
III) Rôles des LT cytotoxiques
• Ils ont un rôle fondamental anti-viral, pour l'élimination des cellules infectées (les virus essayent
de bloquer l'expression du CMH → relai pris par les cellules NK).
• Lutte anti-tumorale.
• Rejet des greffes.
• Activation des macrophages et des cellules NK, par libération de cytokines.
1
/
2
100%