LàOLUTION DES ANIMAUX

1
ANIMAUX
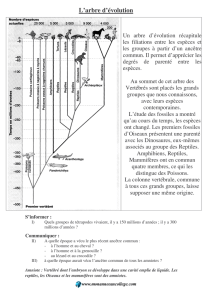
L'ÉVOLUTION DES ANIMAUX
LA VIE AU PRÉCAMBRIEN
La vie apparut sur la Terre pendant le Précambrien ou Archéozoïque, la très longue
période qui s'étend de la formation de la croûte terrestre (il y a 4,6 milliards
d'années) au grand développement des premiers organismes pluricellulaires
complexes (il y a 600 millions d'années). La vie commença il y a environ
3,5 milliards d'années dans l'océan primordial, sous la forme de micro-organismes
très simples : des Bactéries anaérobies qui consommaient les substances
organiques dissoutes dans l'eau. Dans un premier temps, la vie évolua lentement,
jusqu'à ce que, il y a plus de 2 milliards d'années, survienne un grand changement
dans l'atmosphère terrestre : l'activité de certains micro-organismes autotrophes
(appelés Cyanobactéries, ou Algues bleues) suscita l'accumulation d'oxygène et
rendit ce gaz disponible pour d'autres micro-organismes plus actifs et plus
efficaces, les Bactéries aérobies, qui peuplèrent bientôt tous les milieux de la Terre.
En pratique, avec les Cyanobactéries commence la capacité de certains
organismes à fabriquer de la substance organique en libérant de l'oxygène.
L'histoire évolutive des êtres vivants reçut une autre impulsion importante grâce à
l'apparition des premières cellules dotées d'organites internes et d'une membrane
entourant le noyau, il y a environ 1 milliard et demi d'années et, surtout, avec
l’apparition d'une méthode révolutionnaire de reproduction : la reproduction sexuée.
Les Protozoaires, organismes formés d'une seule cellule extrêmement complexe,
représentèrent, il y a environ 800 millions d'années, les premières formes de vie
animale ayant existé sur la Terre. La plupart d'entre eux étaient des
consommateurs qui, pour survivre, avaient donc besoin de se nourrir d'autres êtres
vivants, et qui n'auraient pas pu exister indépendamment des organismes
autotrophes. Le passage suivant fut la pluricellularité, c'est-à-dire le développement
et la diffusion d'organismes formés de plusieurs cellules et ayant des tissus
différenciés pour remplir les différentes fonctions. Les premiers animaux
pluricellulaires (Métazoaires) vivaient dans la mer et étaient complètement
différents de tous les animaux vivants aujourd'hui. Leur corps était mou, mais ils
n'en ont pas moins laissé des empreintes fossiles dans une zone de l'Australie
méridionale, fossiles qui constituent la célèbre faune d'Ediacara. De nombreux
animaux trouvés à Ediacara avaient une symétrie radiale, mais les autres fossiles
présentaient également des traces d'organismes à symétrie bilatérale déjà assez
complexes, dotés de mobilité et de différents organes internes, dont un tube digestif
complet, avec bouche antérieure et anus situés respectivement à l'extrémité
antérieure et à l'extrémité postérieure du corps.
La faune d'Ediacara
La grande différenciation des organismes pluricellulaires au cours du Précambrien
(voir paragraphe précédent) culmina avec la faune d'Ediacara et se termina avec
l'extinction de 98 % des espèces qui avaient évolué jusqu'alors, au point que cette
grande diversification est considérée par les paléontologistes comme un « faux

2
départ » de l'évolution animale. L'extinction eut lieu il y a environ 600 millions
d'années et concerna des animaux complexes à corps mou, ancêtres des actuelles
éponges (Spongiaires), des méduses et des coraux (Cnidaires), ainsi que des vers
marins (Plathelminthes et Annélides), abstraction faite d'autres formes étranges
difficiles à classer. Les fossiles de la faune d'Ediacara témoignent de la
diversification et de la spécialisation atteintes par les premiers organismes
pluricellulaires. Au cours de cette diversification, on eut un développement initial de
tissus, de cellules reproductrices dotées de flagelles et d'organes.
Les Spongiaires et les Cnidaires comme ceux d'Ediacara ont donné en évoluant les
formes modernes d'éponges, de méduses, d'actinies (anémones de mer), de
coraux, etc. Celles-ci se sont différenciées précocement et n'ont pas donné lieu à
d'autres groupes systématiques (phylum) du règne animal. En revanche, les
animaux primitifs à corps plat et symétrie bilatérale constituèrent le point de départ
de l'énorme différenciation d'espèces qui suivit la disparition soudaine de la faune
d'Ediacara et mena graduellement à la formation de la faune actuelle, c'est-à-dire
de tous les phyla de Métazoaires à symétrie bilatérale (Annélides, Mollusques,
Arthropodes, Cordés, etc.) qui peuplent les mers et les terres émergées.
LA VIE AU PALÉOZOÏQUE
Au cours du Paléozoïque ou Primaire, l'ère qui va de moins 600 millions d'années à
moins 250 millions d'années, outre les Invertébrés (éponges, méduses, coraux et
vers), apparurent également dans les mers les Vertébrés.
Le Paléozoïque commença par l'explosion du Cambrien. Il y a 600 millions
d'années, tous les principaux groupes d'Invertébrés encore représentés dans le
règne animal firent leur apparition. Nombre d'entre eux possédaient déjà une cavité
à l'intérieur du corps et quelques-uns présentaient également une segmentation. Un
grand nombre d'espèces appartenant à ces groupes disparut il y a environ
500 millions d'années, laissant toutefois des descendants qui produisirent de
nouvelles formes.
La période suivante, l'Ordovicien (moins 500 à moins 430 millions d'années), se
conclut elle aussi par la disparition de nombreux animaux. L'extinction de
l'Ordovicien fut la première grande extinction de masse dans l'histoire de la vie sur
Terre. Avant cet événement, il y en eut un autre très important : le début de
l'évolution des Vertébrés. Les premières espèces de Poissons primitifs appelés
Ostracodes, apparurent au cours de l'Ordovicien et connurent leur plus grande
expansion pendant le Silurien, la période comprise entre moins 430 et moins
400 millions d'années.
Au cours du Dévonien (de moins 400 à moins 360 millions d'années) apparurent les
premiers Poissons à mâchoires, les Placodermes, apparentés aux formes (les
Acanthodes) qui donnèrent naissance aux Poissons cartilagineux et aux Poissons
osseux, dont la grande diversification eut lieu dans les ères suivantes (voir évolution
des Poissons). Dans la même période, la vie animale commença à s'affirmer sur les
terres émergées. C'est ainsi qu'apparurent les Arthropodes sur la terre ferme, entre
moins 380 et moins 360 millions d'années, et qu'eut lieu l'évolution des Amphibiens,
entre moins 370 et moins 360 millions d'années. L'évolution du Dévonien, qui se
produisit il y a environ 370 millions d'années, intéressa surtout les animaux vivant
dans les mers tropicales.
C'est au Carbonifère (de moins 360 à moins 280 millions d'années), que remonte
l'origine des Reptiles, les premiers véritables Vertébrés terrestres, et des Insectes
ailés.

3
La période suivante, le Permien, qui commença vers moins 280 millions d'années,
se termina par l'événement le plus dramatique de l'histoire de la vie sur la Terre :
l'extinction du Permien, qui eut lieu il y a 240 millions d'années, et au cours de
laquelle disparurent plus de 50 % des animaux qui vivaient dans le milieu marin.
Les Amphibiens furent eux aussi gravement touchés (plus de 90 % des espèces
animales et végétales qui vivaient sur Terre disparurent).
L'explosion du Cambrien
Il y a environ 600 millions d'années, il se produisit une diversification extraordinaire
dans l'évolution des espèces animales, accompagnée de l'apparition de nouvelles
organisations structurelles et de nouveaux modes de vie qui n'avaient pas de
précédent. La plupart des types structurels présents aujourd'hui dans la
documentation fossile firent leur apparition de manière inopinée. Spongiaires et
Cnidaires tout à fait semblables aux formes actuellement vivantes, Mollusques
(Bivalves Lamellibranches ou primitifs), Annélides, Arthropodes (en particulier les
Trilobites) et Échinodermes habitaient, dans une variété d'espèces surprenante, les
mers du Cambrien. Les coquilles et les squelettes externes se développèrent
probablement en réponse à la prédation grandissante due à l’augmentation du
nombre des espèces. Le changement de la composition chimique des océans
rendit disponible une grande quantité de sels nutritifs qui pouvaient être utilisés
pour la formation de squelettes protecteurs. La grande diversification des niches
écologiques fournit les bases de la formation des premières biocénoses complexes
et de chaînes alimentaires.
Comment s'expliquer qu'en à peu près 60 millions d'années soient apparus les
principaux types d'organisation corporelle présents aujourd'hui, tandis qu'aucune
autre nouveauté ne s'est affirmée dans les 500 millions d'années suivantes ?
L'explosion des formes de vie du Cambrien demeure encore un mystère, même si
différentes hypothèses ont été avancées pour l'expliquer. Une des théories de
l’évolution biologique invoque l’idée que l'introduction d'un nouveau niveau (qui
peut, par exemple, être représenté par un prédateur) dans la chaîne alimentaire,
favorise la diversification des espèces aux degrés les plus bas. Par conséquent,
l'apparition d'hétérotrophes pluricellulaires aurait augmenté la diversité des
producteurs, qui à son tour aurait permis la diversification plus poussée des
prédateurs par un mécanisme de rétroaction. Une deuxième hypothèse se rattache
à une théorie plus générale des modalités de l'évolution, dite théorie des équilibres
ponctués, selon laquelle des phénomènes de changement rapide alternent avec de
longues périodes de stagnation. Cette théorie est soutenue de nos jours par de
nombreux paléontologistes et biologistes qui étudient l'évolution, et elle est souvent
confortée par les fossiles que l’on découvre. La souplesse d'un génome assez
simple comme celui des premiers organismes pluricellulaires, surtout au cours du
développement embryonnaire, aurait déterminé une évolution rapide dont les
résultats se seraient « cristallisés » dans les plans corporels présents dans la faune
du Cambrien. La diversification des organismes aurait en outre été facilitée par la
disponibilité de nouvelles niches écologiques. De plus, l'instauration de barrières
géographiques par suite de changements dans la conformation des terres
émergées aurait permis d'orienter dans des directions différentes le cours de
l'évolution dans les différentes zones géographiques.
L'évolution des Poissons

4
Aussi bien les Poissons osseux que les Poissons cartilagineux dérivent
probablement de formes archaïques de Poissons ayant des liens de parenté avec
les Placodermes : les Acanthodes, représentants d'une ligne évolutive qui s'était
détachée précocement d'ancêtres communs avec les premiers. L'appareil
mandibulaire des Placodermes était encore assez grossier, et la mâchoire
s'articulait directement au crâne. L’étape suivante dans l'évolution du squelette qui
entoure la bouche fut celui d'une transformation ultérieure des arcs branchiaux, de
nature à permettre une plus grande mobilité de l'appareil mandibulaire (analogue à
celle d'un requin qui attaque sa proie). Cette nécessité de rendre plus efficaces les
structures impliquées dans la prise de la nourriture eut certainement deux raisons :
l'appauvrissement de la faune des Poissons vers la fin du Paléozoïque et par
conséquent la diversification du régime alimentaire. Les Poissons cartilagineux
(Chondrichtyens) se détachèrent très précocement de la lignée qui mena aux
Poissons osseux (Ostéichtyens), comme en témoignent certaines caractéristiques
fondamentales de leur anatomie (squelette cartilagineux, absence d'opercule
branchial, présence d'excroissances cutanées plutôt que d'écailles osseuses) et de
leur physiologie (mécanisme de régulation de la concentration des liquides internes
fondé sur la rétention de l'urée), et dès le Mésozoïque ils se spécialisèrent comme
prédateurs. Les Poissons osseux apparurent dès le Dévonien, déjà représentés par
deux classes distinctes : les Actinoptérigyens et les Sarcoptérygiens. Les
caractéristiques du crâne et du squelette des nageoires des premiers
Sarcoptérygiens, ainsi que la présence de choanes, rappellent celles des premiers
Amphibiens, qui en sont issus. Des Actinoptérigyens dérivent également tous les
Poissons osseux actuellement vivants. Initialement, il s'agissait de Poissons d'eau
douce ; certains vivaient dans des eaux stagnantes pauvres en oxygène et
soumises à des périodes de sécheresse. Ces conditions portèrent probablement au
développement de « poches » pulmonaires (observables aujourd'hui chez les
Dipneustes). Par la suite, dans les formes qui passèrent dans la mer, ces
« poches » se seraient transformées, pour des exigences hydrostatiques, en vessie
natatoire. La grande diversification des Poissons osseux n'eut lieu que quelques
millions d'années plus tard, au début du Cénozoïque, et les a amenés à devenir
aujourd'hui la classe des Vertébrés qui compte le plus grand nombre d'espèces.
L'évolution des Amphibiens
Entre moins 370 et moins 360 millions d'années, eut lieu l'une des plus
extraordinaires transformations dans l'histoire des animaux. À partir d'un groupe
particulier de Poissons caractérisés par des nageoires robustes au squelette
articulé et des narines communiquant avec la cavité buccale - les Crossoptérygiens
-, l'ancêtre des Vertébrés terrestres d'aujourd'hui conquit la terre ferme. Mais quelle
nécessité avaient les Vertébrés d'abandonner les eaux pour s'aventurer dans un
monde où la force de gravité et le danger de la déshydratation imposaient des
transformations révolutionnaires dans tous les appareils du corps ? La poussée
vers la colonisation des terres émergées fut stimulée par de multiples facteurs.
Dans cette période géologique (vers la fin du Dévonien), le niveau de la mer s'était
abaissé considérablement et l'assèchement partiel de différents bassins contribua
sans doute à un certain nombre d'adaptations à la vie terrestre. En outre, la grande
diversification de la vie marine créait des phénomènes de compétition, les
prédateurs marins s'étaient spécialisés et les proies elles-mêmes étaient très
mobiles. La terre ferme était riche en ressources alimentaires. Quelques dizaines

5
de millions d'années auparavant, en effet, les Arthropodes avaient colonisé les
terres émergées, où désormais les Insectes dominaient les écosystèmes. Ces
Invertébrés constituaient un aliment riche en protéines et, de plus, étaient lents et
donc faciles à capturer. Non pas que les premiers Vertébrés de terre ferme fussent
particulièrement rapides - disons même que leurs mouvements devaient être lents
et gauches -, mais ils étaient plus grands (ils pesaient probablement plus de 10 kg).
Les Vertébrés durent accomplir de nombreuses adaptations avant de s'affranchir
du milieu aquatique. La force de gravité et le danger de déshydratation influèrent
sur la posture et la locomotion à travers l'évolution des membres et d'un squelette
de soutien. La respiration ne pouvait plus se faire à travers les branchies, sujettes à
dessèchement, et c'est ainsi que se développèrent des cavités fermées dites
poumons. Les cellules reproductrices produites par les organes génitaux ne
pouvaient plus être dispersées dans le milieu, et c'est ainsi qu’apparurent la
fécondation interne et les coquilles permettant de limiter la déshydratation des
œufs. Les proies ne pouvaient plus être aspirées avec l'eau (les Poissons et les
Amphibiens primitifs se nourissent de cette manière), mais elles devaient être
saisies : l'évolution de la langue est étroitement associée à cette nécessité. Les
yeux devaient être protégés de la déshydratation au moyen de paupières et de
glandes lacrymales. La différence de densité entre l'air et le corps imposa la
formation de structures (oreille moyenne) capables de transmettre les sons aux
organes de l'ouïe. Ces adaptations mirent très longtemps à se perfectionner et elles
ne sont toujours pas achevées chez les descendants directs des premiers
Vertébrés terrestres, les Amphibiens actuels, qui dépendent encore largement de
l'eau dans leurs cycles vitaux.
Les extinctions du Permien
Entre le Paléozoïque et le Mésozoïque (il y a environ 240 millions d'années) se
produisit l'extinction de masse la plus importante qui ait jamais été enregistrée sur
la Terre. 54 % des familles d'animaux marins et probablement 90 % des espèces
ayant vécu au Permien supérieur disparurent. Cet événement concerna la vie
marine et également une partie de la vie sur la terre ferme. C'est ainsi que
disparurent tous les anciens coraux constructeurs de récifs, de nombreux
Brachiopodes, la plupart des Ectoproctes et des Échinodermes Crinoïdes, les
nummulites (Protozoaires géants munis de coquille, qui se classent parmi les
Foraminifères, atteignant parfois jusqu'à 5 cm de longueur) et de nombreux
Trilobites. Une partie seulement des Vertébrés terrestres subit cette évolution : les
Amphibiens furent touchés dans une large mesure. L’analyse des roches établit
qu'à la fin du Permien la température de la mer augmenta, tout comme l'aridité des
terres émergées. Une fois de plus, les changements climatiques ont été invoqués
pour expliquer les extinctions de masse. Mais comment expliquer cette
augmentation de température ? Un détail important de l'histoire géologique peut
nous aider à faire la lumière sur ces événements. À la fin du Paléozoïque, les
continents se réunirent en un seul supercontinent appelé Pangée (voir passé des
continents), diminuant considérablement l'extension des zones côtières. Si nous
admettons que les zones ayant la faune la plus diversifiée étaient les zones
côtières, et plus généralement celles de la plate-forme continentale, nous devons
en déduire que, quand la Pangée se forma, ces communautés eurent alors un
espace très réduit à leur disposition. L'espace est considéré par les écologistes
comme un facteur limitant la diversité biologique. Moins d'espace, moins d'espèces.
Cette forte limitation de l'espace pour la faune marine côtière, jointe aux probables
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
1
/
71
100%