c-Bilan: Deux MODALITES principales, à l`échelle de l`observation

c-Bilan:
Deux MODALITES principales, à l’échelle de
l’observation géologique (0,25 à 1ma comme degré
maximum de résolution):
- « anagenése » = modifications des caractères au
sein d’une lignée, avec variations des rythmes
évolutifs
-cladogenèse= diversification = multiplication du
nombre d’espèces à partir d’une espèce
originelle
-relation forte entre évolution des caractères et
changements du milieu (sélection naturelle)

Patterns et pas micro - évolution
Gradualisme (Simpson) et équilibres ponctués
(Elredge & Gould)
Gradualisme = changements des caractères de l’ensemble
des populations, progressivement, avec variations de
rythmes (Simpson, etc…)
(nombreux exemples, notamment chez les vertébrés).
Les «sauts»évolutifs correspondent le plus souvent à
des lacunes temporelles de la documentation fossile
2-2- Les mécanismes évolutifs de la spéciation
dans le cadre de l’évolution phylétique: ??

Ponctualisme (=équilibres ponctués, Elredge et
Gould)
= hypothèse développée pour expliquer les « sauts »
évolutifs.
Caractères stables (stases); changements brusques, se
produisant dans une aire marginale de l’aire de répartition
de l’espèce.
L’espèce nouvelle ainsi apparue remplace ou coexiste avec
l’espèce originelle sur la totalité de l’aire de répartition;
ensuite ànouveau stase
(exemples chez les escargots, les trilobites, certains
microfossiles)

En fait, les fossiles ne donnent qu’une
IMAGE du résultat des mécanismes de
l’évolution (pas de temps élevé:
plusieurs centaines de milliers d’années,
des millions de générations…)
Seuls les patterns nous sont accessibles

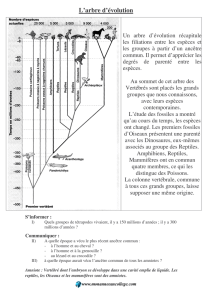
Rappel: Diversification
« explosive » au passage
Précambrien – Cambrien
tous les phyla d’animaux en
quelques millions d’années
= RADIATION EVOLUTIVE
RAZDIATION ADAPTATIVE
3- L’origine des taxons supérieurs à l’espèce:
ordres, classes, phylum
(les « grands groupes »)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%