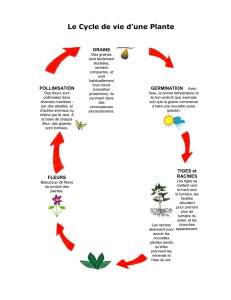
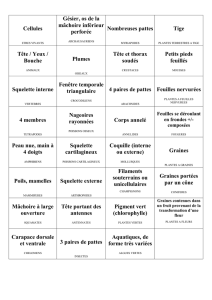
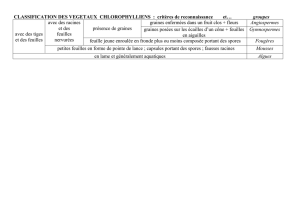
Le potentiel caché du végétal : un allié pour l`humanité

Lilian Ceballos
Le potentiel caché du végétal : un allié pour nourrir l’humanité
Applications agroécologiques pour relever les défis alimentaires,
écologiques et climatiques du XXIème siècle
Edition : Res’OGM Info, Lyon.

Lilian Ceballos
Le potentiel caché du végétal : un allié pour nourrir l’humanité
Applications agro-écologiques pour relever les défis alimentaires,
écologiques et climatiques du XXIème siècle
Introduction
La production alimentaire à l’aube du XXIème siècle
L’agriculture moderne et les questions environnementales et climatiques
La Révolution Verte et ses limites
Les causes du plafonnement des rendements
L’agriculture moderne et ses conséquences
Consensus pour une agriculture durable
Vers une Révolution doublement Verte ?
Au-delà de la productivité
Une approche doublement verte
La transformation des pratiques agricoles
La conquête de la Terre et la diversification des milieux et des interac-
tions
Une brève histoire de la biosphère
La formation de la Terre et l’émergence de la vie
Les vagues successives de pionniers modifient le milieu terrestre
Mise en place des écosystèmes et des interactions durables
Diversification des écosystèmes terrestres et des interactions durables
Vers la complexité du vivant
Les interactions plantes-animaux modèlent les écosystèmes terrestres
Plantes à fleur, pollinisation et dissémination des graines
La Révolution néolithique
La transition chasse-cueillette / agriculture
Origine et diffusion de l’agriculture
Les centres d’origine de l’agriculture

Les fonctionnalités écologiques
Agriculture et artificialisation des milieux
Les fonctionnalités écologiques
Définition des fonctionnalités écologiques
Forçage des fonctionnalités naturelles
Pourquoi les rendements n’augmentent-ils pas indéfiniment avec les doses
croissantes ?
Rôle des fonctionnalités écologiques pour des agro-écosystèmes intensifs et
viables
La régulation du cycle de l’eau
L’épuration des eaux comme fonctionnalité naturelle
La fonctionnalité de structuration et d’ameublissement des sols
La fonctionnalité de fertilité des sols
Les fonctionnalités liées aux traits génétiques de tolérance et de résistance
Les fonctionnalités de compétition pour la lumière et l’espace
Les fonctionnalités liées aux insectes
Les fonctionnalités relatives aux animaux
Les travaux agricoles comme fonctionnalité de l’agro-écosystème
Rôle de la biodiversité comme méta-fonctionnalité
Le potentiel du végétal
Evolution, diversité et fonction des métabolites secondaires
Allocation de ressources à la défense
Evolution des théories de coévolution plantes-herbivores
Multifonctonnalité des métabolites secondaires
Le dilemme des plantes : croître ou se défendre ?
Défense et communication végétale
La localisation des plantes par les insectes
Les défenses indirectes des plantes
Perte du signal dans les génotypes de maïs nord-américains
Communication et induction des défenses
Régulation des gènes de défenses dans les interactions plantes/environnement
Communication et interactions de la plante avec la rhizosphère
Exsudation racinaire et écologie de la rhizosphère
Communication racine/rhizosphère
Communication racine/racine
Communication racine/microbe
Communication racine/insecte
Altération des caractéristiques du sol par l’exsudation

Les mécanismes de l’exsudation racinaire
Localisation spatiale des exsudats racinaires
Communication plantes-bactéries et nodulation
Communication, croissance et signaux chimiques
Développement et défense des nodules
Régulation de la nodulation par les flavonoïdes
Evolution de la spécificité dans la symbiose Légumes-Rhizobium
Modulation de l’activité inductrice par altération de la composition des
exsudats.
Communication plantes-champignons et mycorhization
Diversification et fonction des mycorhizes
Habitat et différentiation fonctionnelle des mycorhizes
Mycorhizes, croissance végétale et nutrition phosphorée
Adaptations racinaires et phosphore
Rôle protecteur des mycorhizes
Les endophytes : entre mutualisme et parasitisme
Fonctionnalités végétales et évolutions morphologiques
L’invention de la graine
La double fécondation des plantes à fleurs
Ecophysiologie de la germination
Influence des conditions environnementales et de la prédation
Intégration des conditions environnementales
La graine comme stratégie de réduction des risques
Influences environnementales sur l’inhibition de la germination
Influence des contraintes biotiques sur la cinétique de germination
Effets de la domestication sur la biologie des graines
Applications agro-écologiques
Optimisation de la capture du rayonnement solaire dans les agro-écosystèmes
Energie lumineuse, photosynthèse et stress oxydatif
Réponse d’acclimatation des végétaux
Cultures associées et semis direct
Cultures intermédiaires et cultures associées pour la nutrition azotée du blé
Impacts des cultures intermédiaires sur le rendement et la teneur en protéines
Les CI améliorent la valorisation de l’azote minéral du sol
Intérêts et limites des cultures associées
Les cultures associées améliorent la teneur en protéines du blé
Pouvoir concurrentiel des variétés de blé
Optimisation du cycle de l’azote
Sources d’azote de l’agro-écosystème

Utilisation des sources d’azote par les plantes
Inoculation de bactéries favorisant la croissance végétale
Association maïs /légumineuses
Fixation azotée et engrais verts
Méthodes de contrôle des ravageurs et pathogènes
Méthodes préventives
La théorie de la trophobiose
Indicateurs locaux et diagnostic
Le respect de la vie sauvage pour bénéficier de l’activité des auxiliaires
Suppressivité des sols
Utilisation d’agents de contrôle biologique
Gestion intégrée des nuisibles
Principes des stratégies de répulsion/attraction (push-pull)
Les composants de la stratégie
Développement des stratégies Répulsion / Attraction
Contrôle des insectes foreurs du mais et sorgho.
Contrôle d’Helicoverpa sur le coton.
Contrôle de Sitona lineatus sur les haricots.
Contrôle du doryphore sur les pommes de terre.
Contrôle des Meligethes du colza.
Contrôle du ver de l’oignon.
Contrôle des thrips sur les chrysanthèmes.
Contrôle des scolytes sur les conifères.
Contrôle de nuisibles en médecine humaine et vétérinaire
Gestion et contrôle des espèces adventices : rôle des rotations et successions
Gestion préventive
Maîtrise des adventices grâce à la succession culturale
Gestion des réserves de graines du sol
Le faux semis
Le désherbage mécanique
Le désherbage thermique
Gestion de la diversité génétique in situ
La graine, organe multifonctionnel
Gestion des flux de gènes sauvages dans les cultures
Le nécessaire équilibre entre dégénérescence et excès de pureté
Régimes et critères de sélection
Conséquences sur l’érosion de la biodiversité
La sélection participative entre amélioration des plantes et sélection paysanne
Le cas du blé : les paysans boulangers
Démarches participatives et variétés maraîchères : les choux
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
1
/
232
100%