Biochimie

- 1 -
Biochimie Franklin et Manue
05 novembre
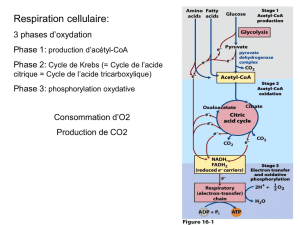
IV- CATABOLISME DU GLUCOSE :
1) Glycolyse :
- Formation d’Acétyl-CoA dans le foie néoglucogénèse.
2) Le shunt des hexoses monophosphates, ou cycle des Pentoses :
1ère voie oxydative irréversible :
- le G6P peut être utilisé dans deux voies différentes :
la glycolyse
le cycle des pentoses
- G6PDH = enzyme de déshydrogénation du -OH en 1chez le G6P, donnant un hémi-acétal,
qui aboutit ensuite à la formation d’une lactone, par l’estérification entre le COOH et OH de
la même molécule (fonction ester interne).
- équilibre entre forme fermée (lactone) et forme ouverte (acide lactonique) acide
gluconique
- Le Coenzyme spécifique de la G6PDH est le NADP.
- 6-Phosphogluconate déshydrogénase : enzyme qui agit en 2 étapes
déshydrogénation, qui aboutit à un produit instable.
décarboxylation, qui aboutit à un sucre à 5 C avec une fonction C=O. C’est le
ribulose-5-P (isomère du ribose-5-P)
- Dans cette réaction, le Coenzyme est le NADP. Il y a donc génération de 1 NADPH pour 1
molécule de 6-phosphogluconate.
- Il y a également dégagement d’une molécule de CO2 par molécule de réactif.
L’ensemble réactionnel G6P ribulose-5-P représente une voie métabolique irréversible, car
il y a perte d’un carbone. Cet ensemble génère donc 2 molécules de NADPH, H+ et 1
molécule de CO2 par molécule de G6P.
2ème voie réversible d’interconversion :
- 3 molécules de ribulose font l’objet de 3 réactions différentes, régies par les enzymes
suivants :
- isomérase : ribulose-5-P ribose-5-P
- épimérase : ribulose-5-P xylulose-5-P (x2)
Ces molécules vont ensuite répondre à des réactions d’interconversion → transcétolase :
ribose-5-P GA3P + sédoheptulose
GA3P + sédoheptulose fructose-6-P + érythrose-4-P
xylulose-5-P + érythrose-4-P fructose-6-P + GA3P

- 2 -
Bilan :
12 NADP+ 12 NADPH, 12 H+
6C6 6C5 = 2C5 + 2C5 + 2C5
6 CO2
2C7 2C3
2C6 2C4 2C6 2C3
2C6
- voir p.10 : On peut considérer que, de manière réversible, on convertit la moitié du GA3P
en DHAP, se traduisant par une réaction dont le fonctionnement est l’inverse de l’aldolase
(2C3 → 1C6).
- 2 GA3P → 1,3 DPG (vers la glycolyse) + DHAP (vers la néoglucogénèse)
- On est parti de 6C6, et on obtient au bilan 2C6 +2C6 + 1C6 (soit 30C). Les 6C restants
correspondent à la production de 6 CO2.
Présentation cyclique de l’ensembles des phénomènes :
12 NADPH
6C6 6C5 - le C6 correspond à du
glucose-6-P
6CO2
C6 5C6
- Les 5C6 associés à un glucose-6-P permettent l’entrée dans le cycle des pentoses → partie
oxydative du cycle.
- En alimentant le cycle d’un G6P, on permet la production de 6 CO2 et 12 NADPH.

- 3 -
- VOIE IRREVERSIBLE : voie oxydative d’irréversibilité conditionnée par la
décarboxylation (4ème réaction).
- VOIE REVERSIBLE : voie dans laquelle on obtient des convertions entre hexoses
monophosphates (G6P et fructose-6-P) et pentoses phosphates (ribose-5-P).
- La voie oxydative fournie la cellule en NADPH, indispensable à un certain nombre de
phénomènes métaboliques :
→ la lipogénèse
→ la synthèse des hormones thyroïdiennes
→ la synthèse de l’ion O2.– par la NADPH oxydase.
→ la synthèse des hormones stéroïdiennes.
→ le maintien du glutathion à l’état réduit.
- La voie réversible à pour rôle de fournir à la cellule le ribose-5-P, qui va permettre la
synthèse des nucléotides et des acides nucléiques.
- Le NADPH protège la cellule des actions oxydantes.
- Le ribose-5-P permet l’anabolisme dans le noyau.
Application pathologique : Déficit en G6PDH.
La voie irréversible est abolie, suite à une mutation du gène. Les cellules vont être
déficitaires dans la production de NADPH, parmi lesquelles les + sensibles sont les globules
rouges.
Du fait d’un déficit en G6PDH, le GR se voit avec un taux réduit de NADPH.
C’est pourquoi ces malades, sous le fait d’agressions pro-oxydantes (aspirine, certains
aliments) souffrent d’une anémie hémolytique.
Le déficit en G6PDH ne limite pas la production de ribose-5-P → l’explication résidant
dans la voie réversible dans laquelle le fructose-6-P reste le précurseur des riboses-5-P.
La cellule peut avoir des besoins réduits en NADPH, et éteindre l’oxydation du fructose
par ces 2 réactions ; mais garder intact la voie d’interconversion pour assurer ses besoins en
ribose-5-P.
Cette pathologie est très répandue sur le continent Africain, parce que les malades
souffrant de ce déficit, sont alors résistants au paludisme (avantage génétique). Le paludisme
est une infection des GR, parasités par le plasmodium. Le déficit en G6PDH fait du GR un
milieu peu propice au développement du plasmodium.
V- LA NEOGLUCOGENESE :
(1, 2 et 3)
- néoglucogénèse = production hépatique de glucose. C’est une voie spécifique au foie.
- Elle est favorisée par l’exercice physique et le jeûne. Elle est perturbée par le diabète.
- Le glucagon et les catécholamines stimulent la néoglucogénèse, alors que l’insuline l’inhibe.

- 4 -
- La néoglucogénèse utilise 3 types de précurseurs :
→ le lactate, libéré par le muscle (mécanisme de CORI)
→ les acides aminés (« glucoformateurs »), provenant du catabolisme des protéines
pour maintenir la glycémie.
→ le glycérol, capté par le foie, vient des TAG, qui sont hydrolysés par des lipases. En
effet, en même temps que l’on est en phase d’hypoglycémie, sont déclenchées
néoglucogénèse et lipolyse.
- La néoglucogénèse lutte en permanence contre l’hypoglycémie.
4) La néoglucogénèse procède selon une voie inverse de la glycolyse, sauf les 3 réactions
irréversibles : l’hexokinase, la PFK1 et la pyruvate kinase.
Pour la néoglucogénèse, ces 3 enzymes sont remplacées par :
→ la G6Pase, spécifiquement localisée dans le foie. Elle hydrolyse la liaison
esterphosphorique.
→ la F(1,6)BPase 1, phosphatase qui hydrolyse en position 1.
Pour le glycérol, il faut 2 autres réactions irréversibles pour arriver au DHAP : la G3PDH et la
glycérokinase.
Le glycérol, capté par le foie, vient des TAG, qui sont hydrolysés par des lipases. En effet, en
même temps que l’on est en phase d’hypoglycémie, sont déclenchées néoglucogénèse et
lipolyse.
NAD+ NADH, H+
Glycérol-3-P DHAP
CH2OH CH2OH
HO H O
CH2O–PO3H– CH2O–PO3H–
Pourquoi la déshydrogénase sélectionnée agit sur le C en 2 (pour donner du DHAP) et non
pas sur le C en 1 (pour donner du GA3P) ?
→ Parce qu’on obtiendrait du L-GA3P. Or, on a besoin de D-GA3P, pour obtenir du
D-glucose.
F(1,6)Bpase G6Pase
2 DHAP → DHAP + GA3P → F(1,6)P2 F6P → G6P glucose
Voir p.11 :
- Lactate et acides aminés : rétroconversion du PU en PEP (réaction irréversible).
- L’implication métabolique se fait dans la mitochondrie et dans le cytoplasme pour assurer la
conversion du Pyruvate en PEP.
- Le foie capte le lactate et le transforme en pyruvate. Le PU est pris en charge normalement
par la PUDH.

- 5 -
Comment le pyruvate fait-il pour revenir, via des réactions dans la mitochondrie, en PEP ?
Nous sommes dans des conditions de lipolyse forte. On a donc un afflux massif
d’acides gras dans les cellules, qui arrivent dans la mitochondrie sous forme d’acétyl-CoA
(suite à la β-oxydation).
- Le cycle de Krebs n’est plus alimenté par le PU mais par l’acétyl-CoA. Le cycle de Krebs
est inhibé car il y a un fort taux d’ATP et de NADH.
Tous les enzymes sont ralentis → accumulation d’acétyl-CoA.
- L’acétyl-CoA inhibe la PUDH, ce qui empêche le PU d’alimenter le cycle de Krebs.
La surproduction d’acétyl-CoA stimule la pyruvate carboxylase, qui permet de contourner la
voie de la PUDH.
PU OA
PU carboxylase
CH3–C–COO– –OOC–CH2–C–COO–
║ ║
O O
+ CO2 + ADP
+ ATP + Pi
- OA évolue vers le malate car on est en présence d’un excès de NADH → déplacement de
l’équilibre de la réaction.
Le malate traverse la membrane mitochondriale (par la navette du malate), d’autant plus
facilement que le malate est aussitôt retransformé en OA. Et ce par oxydation dans le
cytoplasme, car il y a un excès de NAD+, car pas de glycolyse (hypoglycémie).
Dernière étape : la PEPCK permet la synthèse du PEP à partir de l’OA.
PEP
OA OPO3H–
PEPCK Liaison riche en énergie
O=C–CH2–C–COO– CH2=C–COO–
O–
+ CO2
+ GTP + GDP
1
/
5
100%