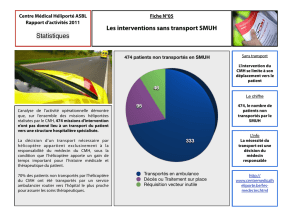
I. Aspect génétique - Cours de DCEM1 2010/2011 à Amiens

Immunologie - E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
COMPLEXE MAJEUR D’HISTOCOMPATIBILITE
- Le complexe majeur d’histocompatibilité (CMH° définit une région chromosomique.
- Identification pour son rôle dans le rejet de greffe chez la souris. Individus génétiquement
identitiques, greffe de l’un sur l’autre été rejetée.
- Chez l’homme on a donné un nom particulier CMH : système HLA (Human Leucocyte Antigen).
Jean Dausset 1958 (Nobel 1980).
- Rôle (explique le rejet de greffe) :
o Présentation antigénique aux lymphocytes T.
o Contrôle de la réactivité des lymphocytes NK.
- Les molécules codées par le CMH sont appelées molécules du CMH (logique). Mais aussi par
extension, on appelle parfois molécules du CMH des molécules impliquées dans la présentation
antigénique, non codées dans le CMH.
I. Aspect génétique
- Gènes situés le chromosome 6.
1. Système multigénique
- Deux classes principales de molécules :
o Classe I (partie 3’) avec les gènes A, B et C.
o Classe II (partie 5’) avec les gènes DP, DQ et DR.
o Classe III : il y a à l’intérieure de nombreux gènes codant pour la réponse immunitaire
mais pas pour la présentation antigénique.
- Les molécules du CMH sont codées par ces gènes A, B, C, DP, DQ, et DR.
2. Système multiallélique
- Polymorphisme : il y a plus 1010 combinaisons possibles à partir de ces systèmes multi-alléliques.

Immunologie - E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
- Le typage HLA d’un individu est quasiment unique.
- Combinaison des différentes allèles sur chromosome est appelée haplotype.
o Puis que les locus classe I, II et III sont intégrés dans une petite région du chromosome,
ces gènes en liaisons étroites ne sont donc pas séparés pendant les mitoses.
o Un haplotype donné est donc transmis en bloc à la descendance.
o Il existe un très faible pourcentage où il y a des recombinaisons possibles entre deux
chromosomes.
- Application clinique importante dans les greffes :
o On regarde quel haplotype l’enfant a reçut de la mère et du père.
o Ainsi on détermine quel membre de la famille est le plus compatible.
- Codominance : chaque allèle est exprimé.
o Groupe HLA : A1 et A2, B8 et B14, etc.
o Certaines HLA sont surexprimées par rapport aux autres. Exemple chez les caucasiens :
HLA2 exprimé par plus de 60% des individus. Donc souvent le même allèle : 2 allèles A2.
II. Structure protéique

Immunologie - E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
- Les gènes HLA A B et C codent pour une chaine lourde comprenant :
o Portion extracellulaire.
o Portion intra-membranaire.
o Portion intra-plasmique.
- Cette chaine lourde s’associe à une chaine légère (non polymorphique pas codé par HLA) β2m.
- Quasiment toutes les cellules nucléées de l’organisme expriment les molécules de classe I.
o Endothélium cornéen : site immuno-privilégié qui n’exprime pas CMH I.
o GR : n’exprime pas CMH I car pas de noyau.
2. Molécules de classe II
- Chaque gène code pour un gène α et un gène β.
- Une protéine de classe de III :
o C’est un hétérodimère formé par une chaine α et chaine β.
o Elle présente une portion membranaire, portion extracellulaire et portion intra-
cytoplasmique.
- Différence classe I et II :
o Les molécules de classe II à l’inverse des classe I sont exprimées sur certains types
cellulaires spécifiques.
o Les cellules qui exprime CMH II sont appelés les cellules présentatrices d’antigène.
o Il existe 3 types qui expriment CMH II de manière constitutive (cellules présentatrices
d’antigène professionnelles): cellules dendritiques, monocytes/macrophages, et LB.
o Types de cellules qui exprime CMH I de manière inductible sous influence des cytokines
(pas spontanée, pas cellules présentatrices d’antigènes professionnelles) : lymphocyte T
et cellules endothéliales.
3. Structure commune
- Le sillon à peptide formé à la surface de la molécule. Le peptide qui peut être accueilli est un
peptide antigénique pour la présentation d’antigènes.
- Conformation dans l’espace similaire :
o Hélice α sur les bordures.
o Sillon formé par un feuillet β plissé. Peptide se loge à l’intérieure du sillon parallèlement
aux hélices α.
4. Différence entre CMH I et CMH II
- Origine différente des peptides présentés par les molécules de classe I et classe II :
o CMH de classe I présente des protéines qui dérivent du cytosol de la cellule : peptides
endogènes (provenant de protéines endogènes à la cellule ou de protéines d’origine
microbienne telle que des protéines virale).
o CMH classe II présent des protéines de l’espace extracellulaire (protéines qui ont
pénétrés dans la cellule par endocytose).
- L’apprêtement de l’antigène (séquence qui assure la présentation d’antigène).
o Dégradation des protéines en peptides.
o Chargement des peptides sur les molécules du CMH préformées :

Immunologie - E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
o Toutes une machinerie est nécessaire pour optimiser la rencontre entre peptides
dégradés et molécules du CMH
- Permet l’expression membranaire des CMH peptides.
- Et la présentation de ces peptides aux lymphocytes T :
o LT CD34+ pour le CMH II.
o LT CD8 pour le CMH I.
- Les résidus d’ancrage permettent la fixation des peptides.
- Le polymorphisme du HLA concerne surtout des résidus d’ancrage :
o Exemple : on s’intéresse au gène HLA A et ses différents allèles (A1, A2, A3, etc.).
o Ces différentes allèles possèdent des résidus d’ancrages différents et fixent donc des
peptides différents.
- Peu de peptides par protéines donnée s’associent à une molécule HLA I ou II donnée.
Si on connait l’haplotype HLA d’un individu on connait les résidus d’ancrage de ces protéines
HLA. Si on s’intéresse à une protéine virale et les différent peptides de sa dégradation, on peut
ainsi caractériser la capacité de chaque individu à faire de la présentation antigénique face à un
virus donné.
- L’allèle HLA Ia est capable de présenté des peptides possédant des acides aminés tyrosine (Y) en
5 et leucine (L) en 8.
- L’allèle HLA Ib est capable de présenté des peptides possédant des acides aminés tyrosine en 2 et
en valine (V) en 9.
- Permet de définir :
o Des petits dominant : forte affinité pour le HLA : qui seront présentés.
o Des peptides sous-dominants : faible affinité pour HLA : peu de présentation.
o Des peptides cryptiques : qui ne s’associent pas au HLA dans des conditions normales.
o Exemple la protéine F du HIV : possèdent des peptides dominants et des peptides sous-
dominants (différent selon l’HLA les individus).
III. CMH de classe I
- En situation physiologique ou pathologique il y aura toujours des peptides présentés sur le CMH
de classe I :
- Présentation en situation physiologie ce sera des protéines endogènes (matrice, métabolisme,
etc.) :
o Des protéines de demi-vie courte.
o Des protéines malformées qui seront dégradées, peptides de dégradation seront
- Présentation en situation pathologie : virus qui produit des protéines virales dans le cytosol.

Immunologie - E. Treiner.
Cours d’Inès Masmoudi. DCEM1 2010-2011.
- Protéines endogène ou protéines virales destinée à la dégradation subissent une ubiquitinylation
par le protéasome.
- Dans le protéasome, des protéines à activité protéasiques clivent la protéine en petits peptides.
- Pendant ce temps dans le réticulum endoplasmique la chaine lourde et la β2m sont synthétisée.
o Elles ne sont pas associées entre-elles.
o Elles sont associées à une protéine chaperonne appelée Calnexine.
- Les peptides entrent dans le réticulum par un système de transport spécifique appelé protéines
TAP (TAP1 et TAP2). Il existe une spécificité selon al longueur des peptides (passent plus ou
moins bien selon la longueur des peptides).
- Rencontre entre CMH I et peptides :
o Fait intervenir un ensemble des protéines chaperones (Erp57 et calréticuline) nécessaires
au chargement des peptides sur les protéines CMH I.
o Il existe une stringence importante concernant les peptides présentés aux molécules
CMH classe I (de 8 ou 9 acides aminés, échelle large mais plus rare 7 à 14).
- Le complexe chaine lourde, β2m et peptide va migrer :
o Par mécanisme d’exocytose formé par appareil golgi.
o Exprimé à la surface cellulaire.
- Protéasome est constitué de 4 x 7 sous-unités protéasiques qui sont :
o Constitutives (existe dans toutes les cellules) car rôle dans la dégradation protéique.
o Inductibles (grâce à certaines cytokines tels que IFN) il y formation d’un immuno-
protéasome. Propriétés un peu différentes en termes de destruction des protéines, il
permet d’améliorer la qualité de peptides présentés aux molécules de CMH classe I :
spécificité augmentée pour le CMH I.
- TAP1 et TAP2 ont une certaine spécificité pour certains peptides en fonction de leur taille :
o Favorise les peptides qui ont la bonne taille pour s’associer au CMH classe I.
 6
7
8
9
6
7
8
9
1
/
9
100%