La vision, phénomène nerveux

1
La vision
A. La vision, phénomène optique
Introduction, quelques observations
. Une grande partie de l’activité humaine dépend de la vision. En absence totale de lumière la vision
devient impossible, cependant certains animaux (Chat, Rapaces nocturnes) peuvent encore voir lorsque
l’intensité lumineuse est très faible.
. Si on essaye de voir simultanément, puis successivement ce qui est écrit au tableau et les empreintes
digitales de ses doigts placés à la hauteur des yeux, on peut faire les constatations suivantes : il est
impossible de voir nettement et simultanément dans deux plans différents ; il faut faire un effort pour
voir un objet proche alors que pour un objet éloigné, on ne ressent aucun effort; l’effort de mise au point
est appelé accommodation.
. On ménage une fenêtre sur la face postérieure d’un oeil de Bœuf, sans léser la membrane mince qu’est
la rétine. (expérience de l’œil décapé). En plaçant un objet lumineux en avant de l’œil, on observe sur la
rétine mise à nu une image renversée, plus petite que l’objet (doc. 2).
I. L’accommodation, phénomène optique essentiel
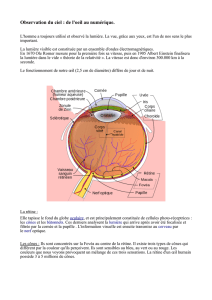
1. L’œil est un appareil d'optique.
.
. La dissection révèle que l’œil est limité par 3 enveloppes emboîtées: la sclérotique (blanche, épaisse et
fibreuse), la choroïde (noire et plus mince), la rétine (peu colorée, mince et fragile) ; sur la face
postérieure de l’œil émerge un cordon blanc, le nerf optique: une coupe longitudinale dans cette région
montre bien que ce nerf est attenant à la rétine en une dépression bien visible appelée papille ou tache
jaune.
. L’œil comprend en outre des milieux transparents de nature à permettre la pénétration de la lumière: la
cornée, l'humeur aqueuse, le cristallin, l'humeur vitrée.
. Le cristallin a une forme de lentille; d'ailleurs si on pose un cristallin frais sur un texte écrit, on constate
qu'il joue le rôle d'une loupe. Il en serait de même pour l'humeur vitrée.
Il est remarquable de constater que cornée, cristallin et humeur vitrée sont des tissus transparents. Plusieurs faits contribuent à leur transparence :
- les cellules qui constituent la cornée et le cristallin ont un contenu transparent.
- L'humeur vitrée (ou corps vitré) est une masse visqueuse, colloïdale ne contenant que de rares cellules.
- La conjonctive, dépendance de la peau, des paupières, est, elle aussi, transparente. D'autre part, à la différence des autres territoires de l’œil, les milieux
transparents ne sont pas vascularisés, ce qui est de nature à favoriser la pénétration de la lumière. Notons que la cornée semble être le seul tissu humain qui
peut être greffé sans réaction de rejet : l'absence de vascularisation est probablement une des causes de cette « heureuse » déficience du système immunitaire.
L'humeur aqueuse n'est pas un tissu, mais du liquide très riche en eau.
L’œil est une suite de
milieux transparents aux
limites desquels (limites
appelées dioptres) les
rayons lumineux
subissent des réfractions
successives (ou
déviations successives).
En plaçant l'humeur
vitrée dans de l'eau, elle
semble disparaître: elle a
le même indice de réfraction que l'eau, soit 1,33. La figure donne les indices de réfraction des milieux
transparents différents .

2
On imagine donc l’œil comme une seule lentille convergente située à 6 mm derrière la cornée, qui aurait
une distance focale de 17 mm et dont le foyer image serait situé dans le plan de la rétine. C’est l’œil
réduit de Lustig, simplifié, donc inexact, mais commode…
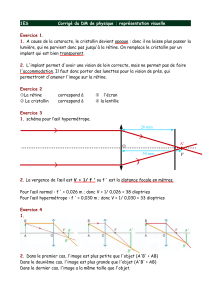
Vergence de l’œil réduit : V=1/f, V= 1/0,017= 58,8, soit + 60 dioptries environ
Dont : 40 pour la cornée, 20 pour le cristallin.
2. Nécessité de l’accommodation
Si on respecte les constructions graphiques ci dessous,
-l’image est nette si l’objet est à l’infini, elle se forme sur la rétine
- pour un objet proche, l’image est floue, elle se forme en arrière de la rétine.
L’accommodation consiste à ramener l’image sur la rétine, elle nécessite un effort de mise au point.
3. Mécanismes de l'accommodation.
a. Hypothèses.
Pour ramener cette image sur la rétine, on peut imaginer plusieurs mécanismes.
. Les différents dioptres oculaires changent de position à la manière des lentilles d'un appareil photo
lorsqu'on réalise la mise au point (appareil à focale fixe) : c'est la distance entre les lentilles et la plaque
photographique qui change.
. La vergence de l’œil varie (appareil à focale variable: zoom) .
b. Analyses des hypothèses: Expérience de Purkinje (fig.3 du poly).
. Dans une salle obscure, on place une bougie allumée latéralement et en avant de l’œil d'un sujet: on
peut observer, sur le fond noir de la pupille, trois images de la bougie obtenues par réflexion .
- Une image droite, très apparente, car très lumineuse, donnée par le dioptre cornéen: I - Une image
droite, peu apparente car peu lumineuse, donnée par le dioptre de la face antérieure du cristallin: II
- Une image renversée, plus petite que la précédente, donnée par le dioptre de la face postérieure du
cristallin: III.
. On invite le sujet à regarder un objet éloigné (à l'infini) , puis un objet proche (ses doigts); pour la vision
proche, l'observation minutieuse révèle :
- l'image I reste inchangée, alors que II devient plus petite (sa taille diminue de moitié si le sujet fixe un
objet situé à une dizaine de centimètres de son oeil) ; l'image III ne diminue pas (ou peu) de hauteur ;
- le diamètre de la pupille diminue ;
- les axes optiques des deux yeux convergent vers l'objet pour le fixer.

3
Ces trois modifications constituent ce qu'on appelle « la triade de fixation », c'est-à-dire les composantes
de l'accommodation. On ne peut pas contrôler ces mécanismes (ce sont des réflexes) ; on estime que
l'accommodation exige un délai d'environ 0,3 seconde.
L'accommodation est donc une déformation de la face antérieure du cristallin: si I est plus petite, c'est que
la face antérieure est d'avantage bombée, donc que son rayon de courbure a diminué.
L'accommodation est donc caractérisée par une diminution de la distance focale du cristallin, donc par
une augmentation de la vergence de l’œil. Le cristallin (donc par conséquence l’œil réduit) est une lentille
à focale variable: sa vergence peut passer de + 20 Dioptries à + 30 à 34D, soit une variation positive de 10
à 14D qui ramène l'image vers l'avant, c'est-à-dire dans le plan rétinien.
c. La déformation du cristallin
. Le cristallin est suspendu en arrière de l'iris par une série de ligaments formant la zonule ; ces ligaments s'insèrent sur une
région différenciée de la choroïde, riche en fibres musculaires, le corps ciliaire formant une couronne autour du cristallin.
. Dans un oeil n'accommodant pas, la zonule est sous tension et le cristallin est donc étiré sur tout son pourtour. Lors de
l'accommodation les fibres musculaires du corps ciliaire se contractent. Ces différentes fibres musculaires agissent de manière
complexe et provoquent le relâchement de la zonule. Libéré de la tension exercée par les ligaments et grâce à son élasticité
naturelle, le cristallin se déforme, se bombe; cette déformation est due au glissement des différentes écailles les unes sur les
autres. Les écailles du centre du cristallin sont plus denses, moins déformables et forment ce qu'on appelle le noyau; le
bombement sera ainsi plus accentué dans la région centrale de cette lentille (son rayon de courbure passe de 11 mm à 5 mm).
d. La diaphragmatisation (fermeture du diaphragme) augmente la profondeur de champ.
e. La convergence des axes optiques permet la formation des images sur les fovéa des deux yeux.
4. Limites de l'accommodation.
. On estime en général que tout objet situé au-delà de 6 mètres est vu sans accommodation: à partir de
cette limite appelée punctum remotum (P.R.) commence l'infini. Les images des objets situés au-delà de
ce point se forment directement dans le plan rétinien (plan focal de l’œil réduit) : l’œil a alors sa vergence
minimale (environ 60 dioptries) ; on dit qu'il est au repos.
. Si nous rapprochons un livre de nos yeux, nous constatons que malgré nos efforts, à partir d'une certaine
distance, les caractères d'imprimerie restent flous: la mise au point est impossible à partir de cette distance
minimale de vision distincte. Cette limite est le punctum proximum (PP. L’œil a alors sa vergence
maximale (74 dioptries) et le cristallin est au maximum de sa déformation.
. L'amplitude d'accommodation est la différence entre la vergence maximale et la vergence minimale: (valeur chez un enfant)
A = V(PP) - V(PR) = 74 - 60 = l4 dioptries
II. Les anomalies optiques de l’oeil (fig. 4 du poly)
On appelle amétropies les anomalies optiques de l’œil.
1. La presbytie.
La distance minimale de vision distincte augmente avec l'âge (voir tableau), ce qui signifie que la
vergence maximale diminue; le presbyte est obligé de lire son journal à bout de bras, mais voit
distinctement des objets éloignés.
Le vieillissement se traduit par une perte d'élasticité du cristallin qui ne peut plus assurer
l'accommodation pour des objets proches: on parle de presbytie lorsque l'amplitude d'accommodation
n'est plus que de 3 à 4. On corrige la presbytie par le port de verres convergents pour la lecture, mais ces
verres ne peuvent pas être utilisés pour la vision de loin. La presbytie peut se combiner avec les autres
défauts optiques de l’œil.

4
2. La myopie.
La myopie est un défaut optique fréquent caractérisé par un P.P. très proche et par l'impossibilité de voir
nettement les objets éloignés (à partir de quelques mètres en général). L'image de ces objets « lointains »
se forme en avant du plan rétinien, ce qui signifie que cette image est floue au niveau de la rétine. La
myopie est un excès de vergence de l'œil dû, soit à l'augmentation de l'indice de réfraction du cristallin,
soit à l'exagération de la courbure cornéenne, soit le plus souvent à l'allongement de l'axe antéro-
postérieur de l'œil. La myopie se corrige par des lentilles divergentes.
3. L 'hypermétropie.
L 'hypermétropie est caractérisée par un P.P. assez éloigné; dans certains cas les images se forment
derrière la rétine même pour l'infini, ce qui oblige les hypermétropes à accommoder sans cesse au prix
d'une fatigue supplémentaire. . . et aboutit à une presbytie plus précoce que chez un sujet normal.
L'œil de l'hypermétrope n'est donc pas assez convergent ou bien l'axe antéro-postérieur est trop court.
Cette anomalie se corrige par le port de verres convergents.
4. L’astigmatisme.
L’image d’un point est une ligne sur certains axes de l’œil; la sphéricité du cristallin ou de la cornée est
imparfaite.

5
La vision, phénomène nerveux
I. STRUCTURE DE LA RÉTINE
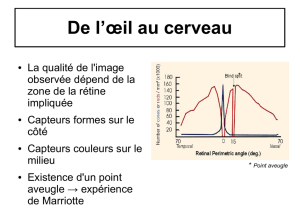
1. La rétine périphérique (fig.5 )
Il s'agit de la partie de la rétine assez éloignée de l'axe par opposition à la rétine centrale située juste dans
l'axe optique.
On distingue nettement dans la rétine, trois couches de noyaux, de densités croissantes en allant de
l'humeur vitrée vers la choroïde. L'embryologie révèle que la rétine est une dépendance de l'encéphale: la
rétine est donc constituée de tissu nerveux, donc surtout de neurones. On peut résumer sa structure en
quatre couches principales :
. Des cellules pigmentaires formant un épithélium accolé à la choroïde ;
. Des cellules très nombreuses dont le prolongement dendritique externe a, soit la forme d'un cône, soit la
forme d'un bâtonnet qui s'insère entre les prolongements de la couche pigmentaire. Les bâtonnets sont
nettement plus nombreux (15 à 20 fois) que les cônes dans la rétine périphérique.
. Des cellules bipolaires moins nombreuses s'articulant par des zones de synapses avec les cellules à cônes
et bâtonnets.
. Des cellules ganglionnaires (appelées encore cellules multipolaires) les plus internes, encore moins
nombreuses, également articulées par une zone de synapses aux cellules bipolaires; les axones de ces
cellules ganglionnaires cheminent à la surface de la rétine pour se réunir à la papille et donner le nerf
optique.
Outre ces trois sortes de neurones, la rétine comprend quelques neurones réunissant horizontalement des
cellules visuelles à d'autres cellules visuelles et les cellules ganglionnaires entre elles. Par ailleurs de
grosses cellules de la névroglie servant de soutien s'y trouvent intercalées. Enfin la coupe microscopique
montre que la rétine contient des vaisseaux sanguins.
2. La rétine centrale
. Du côté temporal, à 3,5 mm de la papille, la rétine humaine présente une zone ovale de 2 à 3 mm,
légèrement colorée et bien vascularisée appelée tache jaune. Cette tache présente en son centre une petite
dépression de 1,2 mm de diamètre : la fovéa.
. Au niveau de la fovéa la rétine présente plusieurs particularités :
- elle ne contient plus de vaisseaux ;
- elle ne contient que des cônes très serrés et de petit diamètre ;
- il y a autant de cellules à cônes que de cellules bipolaires et que de cellules ganglionnaires ;
- les chaînes de neurones sont disposées en gerbe autour de la fovéa ;
- la rétine est nettement amincie à ce niveau.
. Hors de la fovéa le nombre de bâtonnets augmente rapidement.
Conclusion :
La rétine est un tissu nerveux hétérogène dans son épaisseur d'une part et d'une zone à l'autre d'autre part.
Quel est donc l'effet de la lumière sur cette membrane de l’oeil ?
II. NAISSANCE DE L’IMAGE RÉTINIENNE
I. Mise en évidence des propriétés de la rétine.
a. Activité électrique de la rétine.
. A l'obscurité, il existe une différence de potentiel de 6 millivolts entre les faces antérieure et postérieure
Quelques données chiffrées sur la rétine.
Épaisseur: 0,2 à 0,55 mm.
Cellules à bâtonnets: 110 à 130 millions; diamètre : 2 à 5
µ.m.
Cellules à cônes: 6 à 7 millions; diamètre: l,5 µm.
à la fovéa et 4 à 8 . µm dans la rétine périphérique.
Fovéa : diamètre l ,2 mm; 30 000 à 40 000 cônes.
Nerf optique: 500 000 à l 200 000 fibres.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%