biomécanique du muscle strié squelettique

840902276
Fiche de révision – BIOMECANIQUE DEUG 2
PHYSIOLOGIE DU MUSCLE STRIE SQUELETTIQUE
On distingue 3 types de cellules musculaires :
- striées squelettiques
- lisses
- cardiaques
Il existe des différences entre ces cellules au niveau de leur structure, au niveau de leur mode d’activation, au
niveau de leurs propriétés mécaniques, mais les mécanismes de base restent les mêmes.
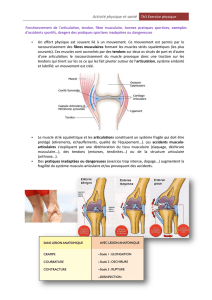
Structure du muscle strié squelettique : (cf. schéma poly)
muscle
faisceaux musculaires enveloppés de tissu conjonctif
fibres musculaires délimitées par le sarcolemme + capillaires sanguins + fibres nerveuses
myofibrilles
sarcomère, délimité par les stries Z, composé de : (bande A + (bande H + (ligne M)) + bande I
filaments épais de myosine constitués de méromyosine légère (tige) + méromyosine lourde (tête
fragments S-1 et S-2)
+ filaments fins d’actine constitué de molécules d’actine + tropomyosine + troponine
fibres nerveuses innervant les muscles = motoneurones A (plus grosses fibres nerveuses de l’organisme)
unité motrice = motoneurone + fibres musculaires innervées par ce motoneurone
rq : Un motoneurone innerve plusieurs fibres musculaires, mais une fibre musculaire n’est innervée que par un
seul motoneurone.
Les propriétés métaboliques et contractiles de la fibre musculaire étant définies par le motoneurone qui
l’innerve, les fibres musculaires d’une même unité motrice seront activées au même instant et de la même
façon.
Le nombre d’unités motrices par muscle dépend de sa taille.
Moins il y a de fibres musculaires par unité motrice, plus le mouvement sera fin.
Mécanisme de la contraction musculaire :
1ere étape : Création d’un PA musculaire.
L’arrivé d’un PA au niveau de l’arborisation terminale d’un motoneurone A provoque la libération de Ca2+.
Ce Ca2+ va provoquer la libération de vésicules d’acétylcholine dans la fente synaptique.
L’acétylcholine va ensuite venir se fixer sur des récepteurs spécifiques intégrés au niveau de la plaque
motrice et ainsi permettre l’ouverture de canaux au Na+.
Le Na+ va pénétrer dans la fibre musculaire, entraînant une dépolarisation de la plaque motrice appelée
potentiel de plaque motrice (PPM).
Cette dépolarisation va se propager de part et d’autre de la jonction neuro-musculaire et créer un potentiel
d’action musculaire.
rq : Le PA d’un motoneurone A est obligatoirement excitateur.
Il existe une certaine durée nécessaire entre 2 PA musculaires car l’acétylcholinestérase, qui est chargée de
dégrader l’acétylcholine, est en nombre beaucoup moins important que l’acétylcholine.

840902276
2e étape : Libération des sites d’actine.
Le PA musculaire se propage le long du sarcolemme, mais aussi au niveau des tubules transverses. La
dépolarisation des tubules transverses va provoquer l’ouverture de canaux calciques et la libération de Ca2+
vers les myofibrilles.
Ce Ca2+ va venir se fixer sur son site spécifique de la troponine et va provoquer la rotation de la double hélice
de tropomyosine, libérant ainsi les sites d’actine sur lesquels vont venir se fixer les têtes de myosine.
3e étape : L’interaction actine-myosine. (théorie des filaments glissants)
Au niveau du fragment S-2 de la tête de myosine, il y a hydrolyse de l’ATP.
La molécule de myosine se charge en énergie chimique et va pouvoir ensuite former un pont riche en énergie
avec l’actine.
L’énergie chimique de ce pont actine-myosine est ensuite transformée en énergie mécanique pour permettre le
déplacement angulaire de la tête de myosine, ce qui rapproche le centre du sarcomère des stries Z et diminue
ainsi la longueur du sarcomère (par raccourcissement des bandes I et H).
3e étape bis : Rupture du pont actine-myosine.
La fixation d’une nouvelle molécule d’ATP va provoquer la rupture du pont actine-myosine. La tête de
myosine reprend sa position initiale et va se retrouver en face d’un autre site d’actine ; un autre pont pourra
ainsi être créé.
4e étape : Le relâchement musculaire.
Le Ca2+ quitte la troponine. La tropomyosine retrouve sa place initiale et bloque les sites d’actine.
Le Ca2+ subit un repompage actif.
Les réponses mécaniques de la fibre musculaire :
La secousse musculaire réponse à un seul PA
- La secousse isométrique : pas de variation de longueur on étudie la variation de tension développée
par la fibre musculaire.
Suite à une stimulation électrique, il faut un attendre un certain temps avant d’observer une tension
musculaire : c’est le temps de latence correspondant au temps de propagation du PA musculaire et au
temps de libération du calcium vers les myofilaments.
Lors du temps de contraction, la tension musculaire croît jusqu’à une valeur maximale.
Lors du temps de relaxation, la tension musculaire retourne vers une valeur nulle ; la durée de
relaxation dépend de la vitesse du repompage du Ca2+.
- La secousse anisométrique isotonique : la tension musculaire est constante on étudie la variation de
longueur de la fibre musculaire.
Le tétanos (en condition isométrique) réponse à un train de PA
On a un phénomène de sommation des secousses. La tension augmente jusqu’à atteindre le palier
tétanique.
Le niveau de tension musculaire dépend de la fréquence des PA.
Les différents types d’unités motrices et de fibres musculaires : (cf. tableau des propriétés des différents types
de fibres musculaires)
3 types d’unités motrices : 3 types de fibres musculaires :
ST I (lipolyse aérobie lentes et résistantes)
FT : - FT IIa (glycolyse rapides et résistantes)
- FF IIb (glycolyse anaérobie rapides et fatigables)

840902276
rq : Les muscles possèdent les 3 types de fibres mais en proportion variable selon le type de muscle (un muscle
impliqué dans la posture aura plus de fibres de type I).
Le recrutement des unités motrices :
Lors d’une contraction progressive :
Recrutement spatial : lorsque la force croît, on recrute de plus en plus d’unités motrices. On recrute
d’abord les unités motrices de types lentes car ce sont les plus petites et donc les plus excitables ; puis
on recrute les FR et ensuite les FF (sauf lors de l’électro-stimulation).
Recrutement temporel : on augmente la fréquence de décharge des PA jusqu’à atteindre le tétanos
parfait.
rq : A 80% de la force maximale, toutes les unités motrices sont recrutées. Au-delà, c’est la capacité de
recrutement des unités motrices rapides qui joue.
La comparaison de la force maximale développée par un muscle activé électriquement avec la force
maximale développée volontairement par un sujet permet d’évaluer sa capacité d’activation volontaire
de ses muscles.
Lors de différentes activités :
posture 25% d’unités motrices recrutées
marche 50% d’unités motrices recrutées
Plus l’intensité de l’exercice augmente, plus il y aura d’unités motrices recrutées.
Plasticité des unités motrices :
Myotypologie = composition d’un muscle en type d’unités motrices.
fixée à 70% par le code génétique
Toutes les conversions de fibres sont possibles (y compris de lentes à rapides).
Il faut jouer sur les propriétés des motoneurones : conduction de l’influx nerveux (épaisseur de la gaine de
myéline), synthèse des neuromédiateurs, vitesse de transmission synaptique.
rq : Les propriétés des motoneurones déterminent les propriétés des fibres musculaires en kinésithérapie, on
aura donc d’abord une réadaptation neuromusculaire avant la musculation.
Lors de mouvements pliométriques, les muscles à contraction lente ont tendance à devenir rapides.
BIOMECANIQUE DU MUSCLE STRIE SQUELETTIQUE
Les différents types de contraction :
- isométrique : pas de variation de longueur (Fm = C)
- anisométrique concentrique : raccourcissement du muscle (Fm > C)
- anisométrique excentrique : allongement du muscle (Fm < C)
(- pliométrique)
Si Fm = constante, la contraction est isotonique.
Si Fm varie, la contraction est anisotonique.

840902276
rq : Si l’on travaille un muscle dans un seul type de contraction, on a un gain de force uniquement dans ce type
de contraction il faut donc travailler dans tous les types de contraction.
Section physiologique = nombre de filaments dans un muscle
détermine la tension musculaire maximale.
dépend du type de muscle : un muscle fusiforme, plutôt impliqué dans un mouvement dynamique, est moins
puissant qu’un muscle penniforme, plutôt impliqué dans la posture.
Modèle de HILL :
Schéma :
C.E.S.
C.C.
C.E.P.
Composante Contractile (C.C.) : c’est le générateur de force, le lieu où est transformée l’énergie
chimique en énergie mécanique (au niveau des ponts actine-myosine).
Relation force – longueur :
Schéma expérimental :
Courbe :
Il n’est pas possible de mesurer la tension active (relative à la CC) développée par le muscle, il faut donc
la calculer grâce à la relation : tension active = tension globale - tension passive (relative à la CEP)

840902276
Lorsque l l0, on a une tension musculaire maximale.
Lorsque l > l0, on a un allongement du muscle. La tension musculaire diminue jusqu’à être nulle pour
175% de l0.
Lorsque l < l0, on a un raccourcissement du muscle. La tension musculaire diminue jusqu’à être nulle
pour 60% de l0.
Cette allure parabolique de la relation tension active – longueur s’explique par la théorie des filaments
glissants : la tension active dépend du degré de chevauchement des filaments d’actine sur les filaments
de myosine.
Schéma :
Relation force – vitesse :
Schéma expérimental :
Courbes :
Courbe : Relation force – vitesse
 6
7
8
9
6
7
8
9
1
/
9
100%