CM PHYSIO n°1

CM PHYSIO n°1
Le 7/10/1998
INTRODUCTION
OBJECTIF : Connaître le fonctionnement des grandes fonctions de l’organisme
dans le but de comprendre les adaptations physiologiques à l’exercice.

Chapitre I
NOTION DE CHIMIE
ET DE BIOCHIMIE
I L’atome :
C’est le plus petit niveau d’organisation des êtres vivants. Son unité de mesure est
l’Angström.
1 A = 10-10m
Rappel : 1 nm (nanomètre) = 10-9m 1 m (micromètre) = 10-6m
Les atomes sont constitués de trois éléments : électrons, protons et neutrons.
Particules infra-atomiques
Particules
Charge
Masse relative
Electron
Négative
1
Proton
Positive
1836
Neutron
Neutre
1838
On considère que proton et neutron ont la même masse relative.
D’après ce tableau, le neutron est 1838 fois plus lourd que l’électron.
Les électrons gravitent car ils sont attirés par les forces positives.
Dans un atome, il y a autant de protons que d’électrons, ce qui signifie que
l’atome est électriquement neutre.
Nombre atomique : C’est le nombre d’électrons qui tournent autour du noyau.
Poids atomique : C’est la somme du nombre de protons et de neutrons.
Comme les atomes sont très petits, on utilise une unité de quantité particulière : la
mole (M ou mole) qui contient 6,02 1023 éléments.
On sait qu’un atome de carbone C pèse 12 fois plus qu’un atome d’hydrogène. Comme
une mole contient toujours le même nombre d’éléments : une mole d’atome de C pèse
12 fois plus qu’une mole d’atome H, soit 12g.
La concentration, c’est la qtité de matière par unité de volume (en g ou mmol/L )
Résumé :
L’atome est constitué de 3 types de particules qui diffèrent par leur charge électrique et
leur masse relative.

Les protons qui portent une charge positive forment avec les neutrons, particules non
chargées, le noyau de l’atome et lui confèrent une charge positive. Des électrons
chargés négativement, attirés par la charge positive du noyau gravitent autour de ce
noyau.
II Les molécules :
La réunion d’atomes par des liaisons chimiques aboutit à la formation de molécules.
Pour qu’il y ait liaison chimique, les atomes doivent avoir en commun au moins un
électron.
ATOME
NOMBRE DE LIAISONS POSSIBLES
H
1
C
4
N
3
O
2
III Ions et molécules polaires
L’électron de la molécule avec le moins de protons est attiré par la molécule avec le
plus de protons.
Ex : H2O H+
Un atome a tendance à perdre un électron, il est
O électropositif.
H+
La liaison chimique entre atomes peut être de différents types (neutre ou polaire).
La perte d’un électron par un atome donne naissance à un ion positif (cation) et le gain
d’un électron donne naissance à un ion négatif (anion).
Selon le type de liaisons chimiques à l’intérieur d’une molécule, celle-ci peut être
neutre, polaire ou ionisée.
Les forces électriques assurant la cohérence de la molécule sont source d’énergie
potentielle qui peut être libérée à l’occasion de la rupture de ces liaisons.
IV Les états de la matière
Solide, liquide, gazeux : il existe 3 états de la matière. Dans les états de la matière, les
molécules qui constituent la matière sont en perpétuel mouvement.

A l’état solide, le mouvement des molécules est réduit car l’attraction entre les
molécules est très forte.
A l’état gazeux, l’attraction entre les molécules est moins forte, le mouvement des
molécules est très grand.
A l’état liquide, état intermédiaire entre les 2.
En augmentant la température, on augmente le mouvement des molécules, car elles
libèrent des forces d’attraction.
Un liquide dans lequel se dissout une molécule s’appelle un solvant.
Pour qu’un solvant puisse dissoudre, il faut qu’il y ait interaction entre les
molécules.
V Classification des molécules présentes dans le vivant
Il existe deux grandes familles de molécules :
- l’eau et les sels minéraux, ils constituent les substances inorganiques car
elles ne possèdent pas d’atome de carbone.
- les substances organiques, comprenant 1 ou plusieurs atomes de carbone.
Composition de l’organisme en molécules
Constituant
Fraction de la masse brute, en %
Eau
Protéines
Lipides
Minéraux
Corps intermédiaires et acides nucléiques
Glucides
60
17
15
5
2
1
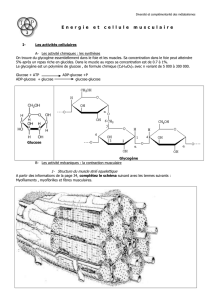
1° ) Les glucides (les oses) :
Ce sont les molécules les plus simples de l’organisme car leur nombre de carbones
est compris entre 3 et 7.
Ces atomes de carbone ne portent que des atomes d’oxygène ou d’hydrogène.
les trioses ( 3 atomes de carbone ) : ex : glycéraldéhyde qui constitue un
carrefour essentiel du métabolisme cellulaire (p.4 )
les pentoses (5 atomes de carbone ) ex : ribose nécessaire à l’élaboration de
plusieurs molécules comme l’ADN (p.4 )
les hexoses (6 atomes de carbone ) : ex : glucose, fructose ou galactose qui
sont des molécules alimentaires essentielles pour la cellule (p.4 )

C’est l’alimentation qui nous apporte les glucides nécessaires au fonctionnement de
l’organisme, notamment dans le lait qui contient des monosaccharides. La majorité
des sucres proviennent de l’alimentation d’origine animale.
Le rôle des glucides est de constituer une réserve énergétique pour nos cellules.
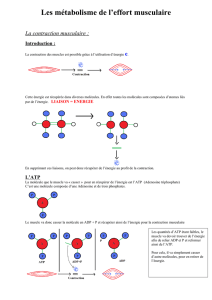
C’est grâce à la dégradation du glucose que l’on peut fabriquer de l’énergie
indispensable à la contraction musculaire.
Quand le glucose n’est pas utilisé sous forme d’énergie, il est stocké sous forme
de glycogène.
Lorsque l’apport alimentaire en glucose est insuffisant, les cellules hépatiques
synthétisent du glucose. Le glucose excédentaire est stocké sous forme de graisse
dans des cellules spécialisées qui constituent le tissu adipeux.
Les sucres ont un rôle de substrat énergétique de manière à fabriquer de l’énergie.
Dans certaines conditions, le foie peut transformer le glucose en acide aminé, qui
sont les principaux constituants des protéines.
2° ) Les lipides
Ils possèdent très peu de groupements polaires ou ionisés, ils sont donc insolubles
dans l’eau.
Ils existent sous deux états : solide, on les appelle des graisses et liquide, ce sont
alors des huiles. En fonction de leur structure, ils se répartissent en 3 groupes :
Les graisses neutres (p.4)
On y retrouve les acides gras : par exemple, l’acide palmitique.
Triglycéride : Réserve la plus concentrée de l’organisme. Forme sous laquelle les
acides gras sont stockés dans l’organisme. Il est possible de dégrader des
triglycérides pour former de l’énergie.
Les triglycérides sont stockés dans des cellules qui constituent le tissu adipeux :
les adipocytes. Le tissu adipeux forme un coussin protecteur autour des organes.
Les phospholipides (p.6)
Ils sont polarisés en raison du groupement phosphate.
Dans l’eau, les phospholipides s’orientent car ils possèdent une «tête» polaire et une
« queue » non polaire.
Les parties non polaires s’orientent vers les parties lipidiques et les parties polaires
vers les parties aqueuses.
Les phospholipides sont les principaux constituants des membranes cellulaires
et des gaines de myéline.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
1
/
81
100%