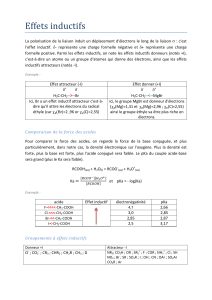
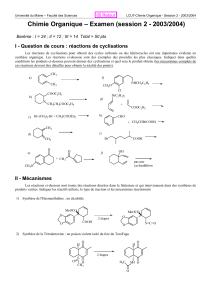
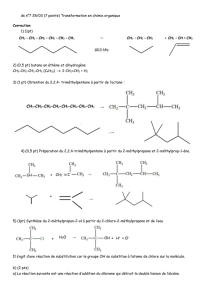
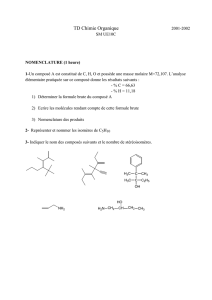
9 metabolisme hemoglobine

UEHIN - Ecole de Médecine Saint Christopher Iba Mar Diop - Métabolisme de l’Hémoglobine – Octobre 2012
1
METABOLISME DE L’HEMOGLOBINE
I. GENERALITES
L’hémoglobine est une hétéroprotéine du sang, responsable du transport de l’oxygène des poumons
vers les tissus d’une part, et d’autre part le transport du gaz carbonique des tissus vers les poumons.
L’hémoglobine représente environ 35 % de la masse des hématies dans lesquelles elle est contenue.
L’oxygène circule sous forme majoritairement liée à l’hémoglobine. En effet 98 % de l’oxygène
total contenu dans le sang est fixé à l’hémoglobine.
II. BIOSYNTHESE DE L’HEMOGLOBINE
II.1. BIOSYNTHESE DE L’HEME
L’hème est la porphyrine des hémoglobines, qui va permettre la fixation de l’hémoglobine grâce à
son atome de fer central. De façon générale, les porphyrines sont des corps cycliques formés de
quatre anneaux pyrroliques et dont l’une des principales propriétés est de former des complexes
avec les ions métalliques (hème, cytochrome, catalase, chlorophylle…)
La biosynthèse de l’hème s’effectue en plusieurs étapes, alternativement dans la mitochondrie et
dans le cytosol (figure 1).
Figure 1 : Localisations subcellulaires des étapes de la biosynthèse de l’hème (GRANICK)
Formation du succinyl CoA
Succinate + GTP + CoASH Succinyl CoA + GDP + Pi
Succinate thiokinase
L’acide succinique provient du cycle de Krebs, d’où la nécessité de fonctionnement optimal de ce
cycle pour alimenter la cellule en succinate. Le succinyl CoA peut également provenir du cycle de
Krebs.
Formation de l’acide delta amino lévulinique (AAL)
+
COOH
CH2
CH2
CO SCoA
Succinyl CoA
Glycine
CH2
COOH
NH2
CO2
CoA-SH COOH
CH2
CH2
C
O
CH2 NH2
Acide -amino lévulinique (AAL)
COOH
CH2
CH2
C
O
CH NH2
COOH
Acide -amino -cétoadipique
Figure 2 : Synthèse de l’acide delta-amino lévulinique (AAL)

UEHIN - Ecole de Médecine Saint Christopher Iba Mar Diop - Métabolisme de l’Hémoglobine – Octobre 2012
2
Cette réaction se déroule dans la mitochondrie en présence d’aminolévulinate synthase (figure 2).
Cette enzyme allostérique mitochondriale fonctionne avec le phosphate de pyridoxal (PAL) et est
rétroinhibée par l’hémoglobine, l’hème et la protoporphyrine IX.
Formation du porphobilinogène (PBG)
Cette réaction est catalysée par la porphobilinogène synthase liée à la phase aqueuse cellulaire. Il en
résulte une condensation de deux molécules d’AAL suite à deux déshydratations internes, d’où le
nom de AAL déshydratase également donné à l’enzyme catalysant cette réaction (figure 3).
2H2O
CH2
NH2
O
COOH
CH2
CH2
C
COOH
CH2
CH2
C
O
CH2
H2N
C
CH2
NH2
COOH
CH2
CH2
CH
HN
COOH
CH2
C
C
AAL + AAL Porphobilinogène (PBG)
Figure 3 : Formation du porphobilinogène (PBG)
Formation des dérivés tétrapyrroliques (porphyrines)
La formation d’un dérivé tétrapyrrolique se fait grâce à la condensation de …………………
La condensation se fait grâce aux carbones porteurs de la fonction aminée (carbones de la
glycine). Ces carbones deviennent alors les carbones méthyléniques et qui relient les
quatre dérivés monopyrroliques en un seul dérivé ……………………………………………
La réaction est catalysée par la porphobilinogène désaminase.
- Le premier dérivé tétrapyrrolique obtenu est l’uroporphyrinogène (UPG) qui possède quatre
isomères possibles, mais dont deux seuls sont présents dans la nature : l’isomère I (0,1 %) et
l’isomère III (99,9 %). Parmi ces deux, c’est l’isomère III (UPG-III) qui est retrouvé dans le
.métabolisme de l’hémoglobine (figure 4).

UEHIN - Ecole de Médecine Saint Christopher Iba Mar Diop - Métabolisme de l’Hémoglobine – Octobre 2012
3
Acétate (A)
Propionate (P)
(A)
(P)
(A)
(P)
(P)
(A)
A P
A
P
P A
P
A
C
N
H
C
C
C
CCH2
H2
HOOCCH2
COOH
CH2
H2C
C
C
HN
CC
CCH2
CH2
COOH
CH2COOH
C
H
N
C
CC
CH2
CH2
HOO
C
CH2
H2CCH2
COOH
C
C
C
C
NH
C
C
C
CH2
COOH
H2CH2CCOOH
A
P
A P
A
P
P A
UPG-I UPG-III
Figure 4 : Structure de l’uroporphyrinogène III (UPG-III)
- L’UPG-III se transforme en coproporphyrinogène III (CPG-III) par décarboxylation de tous
les groupements acétate (A) qui deviennent des groupements méthyle (M). Cette réaction est
catalysée par l’uroporphyrinogène décarboxylase (figure 5).
A
P
A P
A
P
P A
UPG-III
M
P
M P
M
P
P M
CPG-III
CO2 (4)
Figure 5 : Passage de l’uroporphyrinogène III au coproporphyrinogène III
- Le CPG retourne dans la mitochondrie où il est transformé en protoporphyrinogène III
(PPG-III) puis en protoporphyrine III. La coproporphyrinogène oxydase catalyse la
décarboxylation et l’oxydation de deux chaînes latérales propioniques (P) qui se
transforment en groupements vinyles (V) et donner ainsi le protoporphyrinogène III.
- Secondairement, le PPG est oxydé en protoporphyrine grâce à la protoporphyrinogène
oxydase (figure 6 et 7).

UEHIN - Ecole de Médecine Saint Christopher Iba Mar Diop - Métabolisme de l’Hémoglobine – Octobre 2012
4
M
P
M P
M
P
P M
CPG-III
M
P
M V
M
V
P M
PPG-III (IX)
O22 CO2
M
P
M V
M
V
P M
Protoporphyrine-III (IX)
6 H
Figure 6 : Passage du CPG au PPG et la Protoporphyrine IX
CH
CHHC
HC N
HN
N
NH
M V
M
V
P M
M
P
Figure 7 : Structure de la protoporphyrine IX
Formation de l’hème
La synthèse de l’hème est parachevée dans la mitochondrie par l’incorporation de fer ferreux au
noyau protoporphyrine IX grâce à l’hème synthase ou ferrochélatase.
Régulation
Le point de régulation clé est constitué par la réaction catalysée par l’AAL synthase qui est une
enzyme allostérique.
II.2. BIOSYNTHESE DE LA GLOBINE (Voir synthèse protéique)
II.3. LIAISON HEME-GLOBINE
Cette union se déroule dans le cytosol. Chacune des quatre chaînes de la globine fixe un hème par
l’intermédiaire d’un histidine qui fournit un doublet électronique d’un azote de son cycle. C’est la
5ème liaison offerte au fer, les quatre premières l’étant avec les quatre azotes des quatre cycles
tétrapyrroliques. Une sixième liaison est contractée lors de la fixation de l’oxygène (figure 8).

UEHIN - Ecole de Médecine Saint Christopher Iba Mar Diop - Métabolisme de l’Hémoglobine – Octobre 2012
5
Fe
HIS (63)
NO2
Figure 8 : Liaison Hème-Globine-Oxygène
III. CATABOLISME DE L’HEMOGLOBINE
Les érythrocytes sénescents sont éliminés de la circulation générale et catabolisés au niveau du
système réticuloendothélial.
La partie protéique de l’hémoglobine est hydrolysée en ses acides aminés constitutifs, tandis que la
partie héminique va suivre des voies complexes de dégradation donnant des intermédiaires
communément appelés pigments biliaires (figure 9).
Ces pigments sont caractérisés par l’ouverture oxydative du cycle tétrapyrrolique entre les cycles I
et II. On obtient alors une chaîne linéaire tétrapyrrolique appelée biliverdine de coloration verdâtre.
La biliverdine est secondairement transformée en bilirubine grâce à la biliverdine réductase. La
bilirubine est un pigment biliaire jaune toxique pour l’organisme car est en mesure de découpler la
respiration. La bilirubine est ensuite amenée par voie sanguine jusqu’au foie sous forme d’un
complexe avec la serumalbumine.
HEME(
CH
CHHC
HC N
N
N
N
M V
M
V
P M
M
P
I
II
III
IV Fe3+
O2
Fe3+
CO
 6
7
8
9
6
7
8
9
1
/
9
100%