I. La contraction musculaire

Physiologie Générale Pr. Tobias Hevor
1
Physiologie animale
CH1 : Les cellules nerveuses
I. Découverte de la neurobiologie
La physiologie = étude du fonctionnement du tissu, des organes, des cellules entières et des
organismes entier.
1865 Jan PURKINJE (tchèque) = découverte des cellules de PURKINJE
1865 Otto DIETERS (Bonn) = découverte des dendrites et des axones
A. Camillo GOLGI = utilise la technique de golgi (utilisation de nitrate d’argent) pour observer
les cellules nerveuses
1888 Santiago RAMON = identifie les neurones
1887 Wilhem HIS
1888 August FOREL
1891 Wilhem WALDEYER
Un neurone = une cellule capable d’émettre un message, de propager un message, de communiquer
ce message a une autre cellule. Les neurones ont aussi la capacité de sécréter des substances et ont
donc une fonction sécrétrice.
Plusieurs neurones forment un circuit neuronique.
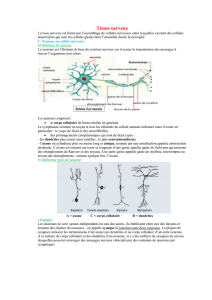
II. Cytologie du neurone
A. Aspect général
La membrane des neurones est en mosaïque de type SINGER NICHOLSON (lipide + protéine+ protéine
particulière).
A l’extérieur de cette membrane on retrouve :
- des fibronectines
- des protéines d’adhésion (Cell Adhesion Molecule = CAM => neural-CAM = N-CAM)
A l’intérieur :
- de l’actine (tension du neurone)
- du NICHOLSON (tension du neurone)
- de la spectrine (formation de la forme du neurone)
- de la fodrine (formation de la forme du neurone)
Le noyau des neurones est « spécial » en effet :
- il n’y a plus de mitose dans les neurones = noyau a phase de repos permanent
- la chromatine est uniforme
- le nucléole a une taille importante (= fabrication de protéine importante)
- présence d’organes circumventriculaire
- les pores nucléaires sont en grand nombre
- présence de petites molécules = stéroïdes

Physiologie Générale Pr. Tobias Hevor
2
Le réticulum endoplasmique (RE) est très actif, est s’appelle le corps de NISSL car très développé. De
plus les lysosomes sont très fonctionnels, les mitochondries sont très actives et le cytosquelette de la
cellule est très développé
Les protéines synthétisées peuvent être classées selon 3 familles :
- les protéines qui reste dans le cytoplasme = gel cytoplasmique
- les protéines qui sont incorporées dans les différents organites (noyau, mitochondrie,…)
- les protéines véhiculées le long des prolongements (dendritiques, récepteur) = servent à
renouveler la membrane des axones.
B. Le corps cellulaire et les dendrites
Corps cellulaire = SOMA = péricaryon.
Le SOMA est associé d’un coté aux dendrites (pole récepteur du neurone ou les informations arrivent)
et de l’autre coté a un axone (pole émetteur du neurone qui transmet les informations).
Le corps cellulaires est polymorphe c'est-à-dire qu’il a des formes variables :
- les cellules pyramidales = hippocampe
- les cellules ovoïdes (en œuf) = cellules de PURKINJE
- les cellules fusiformes = pallidum

Physiologie Générale Pr. Tobias Hevor
3
- les cellules étoilées = moelle épinière
- les neurones en T = ganglion de la moelle épinière. Ce sont des cellules sensorielles qui n’ont
pas de dendrites
- les neurones bipolaires = rétine.

Physiologie Générale Pr. Tobias Hevor
4
C’est le SOMA qui assure l’intégrité de la cellule du neurone et par conséquent qui fabrique les macro-
protéines nécessaire à la régénération cellulaire.
Les dendrites sont des extensions du cytoplasme, il y en a environ 9 lorsque l’on quitte le corps
cellulaire mais elles sont recouvertes d’épines dendritiques (environ 10000 épines par dendrite),
cependant certains neurones sont lisse et n’ont pas d’épines dendritiques. Les dendrites ont un
diamètre qui diminue au fur et à mesure que l’on s’éloigne du corps cellulaire.
C. L’axone et les collatéral
L’axone est toujours lisse et capable d’avoir des ponts collatéraux, ce pont collatéral peut revenir sur
son propre SOMA= pont collatéral récurent.
L’axone est recouvert par une gaine isolante = gaine de myéline.
Les neurones de type Golgi I = neurone cortico-thalamique (neurones assez long).
Les neurones de type Golgi II = neurone permettant la création de circuit locaux (neurone
relativement court).
L’axone se ramifie à son extrémité, chaque extrémité va se renfler en bouton (bouton terminal). Un
bouton + un récepteur forme une jonction plus communément appelé synapse.
Le pole collatéral est considéré comme le pole d’émission. Au niveau du segment initial il y a la
création d’un potentiel d’action qui se propage le long de l’axone et permet la libération de substance
(neurotransmetteur) qui permet la transmission du message. Ce message peut être modulé au niveau
du segment initial.
III. Synthèse et trafic intra neuroniques
A. Maintient de la forme des neurones
Le cytosquelette est le support matériel :
- microtubule (25 nm de diamètre)

Physiologie Générale Pr. Tobias Hevor
5
- les Microtubule Associated Proteine ou MAPs = charpente de l’axone
- l’actine globulaire (actine G)
Il faut deux filaments d’actine F pour faire un filament d’actine
- des filaments intermédiaires (7 à 11 nm), ce sont des neurofilaments constitués de 3 protéines
agglomérées.
Les microtubules/filaments sont des éléments labiles (qui ne sont pas fixe) qui se polymérisent plus du
coté axonique que du coté dendritique. Le transport de ces microtubules est rétrograde et
antérograde.
B. Synthèse de macromolécules
Les mitochondries sont allongées. Dans les dendrites se trouvent le RE et les polysomes mais l’appareil
de golgi ne se trouve pas dans les dendrites. Dans les dendrites se trouvent des MAPs spécifiques aux
dendrites.
Les ARN sont transportés du corps cellulaires jusqu’aux dendrites (500μm par jour) ou sont synthétisé
des molécules de petites tailles.
Dans l’axoplasme il n’y a ni RE ni appareil de golgi, il n’y a donc pas de possibilité de synthétiser des
protéines dans l’axone. Les protéines de l’axoplasme sont donc synthétisées dans le SOMA.
IV. Transport axonique
A. Mise en évidence
Pour mettre en évidence ce transport axonique on utilise des axones géants de cellule de calmar qui
font environ 1 mm de diamètre.
Expérience : on pose une ligature sur l’axone de calmar et on observe un renflement juste avant la
ligature.
On peut donc en conclure qu’à l’intérieur de l’axone du matériel a été bloqué par la ligature.
Néanmoins si on met une autre ligature on observe un autre renflement de l’autre coté de la ligature.
Il y a donc un transport de matière dans les deux sens :
- transport antérograde/extracellulaire = noyau queue
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
1
/
35
100%