CHAPITRE III CLIVAGE - BLASTULATION

CHAPITRE III CLIVAGE - BLASTULATION - GASTRULATION
3. 1. Généralités
La fécondation mène à la constitution d'un nouvel organisme mais active aussi l'ovule
qui, devenu zygote, entame immédiatement sa première mitose. L'embryon passe ainsi à l'état
pluricellulaire. Les premières mitoses se succèdent à l'intérieur de la zone pellucide, sans
modification significative de la taille de l'embryon. Les cellules filles ou blastomères
deviennent ainsi de plus en plus petites et voient leur rapport nucléo-cytoplasmique
augmenter. Elles demeurent cependant indépendantes les unes des autres. C'est la phase de
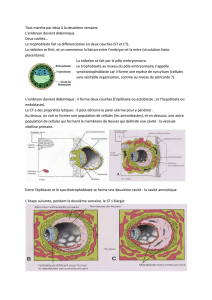
clivage ou segmentation (figure 3.1). Dès la deuxième division (passage du stade 2 cellules
au stade 4 cellules), les divisions sont asynchrones. Chez les vertébrés, la morphologie de la
segmentation varie suivant le type d'ovule (cf. chap. II).
Chez les mammifères euthériens, le stade suivant est caractérisé par la mise en place
de jonctions intercellulaires qui solidarisent les blastomères et modifient la morphologie de
l'embryon pour le faire ressembler à une mûre : c'est le stade morula (figure 3.1).
Les divisions se poursuivent et, au sein de ce massif compact de cellules, apparaît une
cavité, le blastocèle (ou blastocœle). Cette cavité caractérise le stade blastula ou blastocyste.
L'embryon est alors constitué d'une assise externe de cellules épithélioïdes qui limitent la
cavité, le trophoblaste ou trophectoderme, auquel est appendu un petit massif de cellules non
différenciées, la masse cellulaire interne ou bouton embryonnaire (figure 3.2). Le stade
blastocyste est donc le siège du premier processus de différenciation cellulaire. Le
trophectoderme peut être considéré comme la première annexe extraembryonnaire et tiendra
un rôle important dans l'implantation. La masse cellulaire interne fournira la totalité du fœtus
et l'essentiel des annexes extraembryonnaires. Sous la pression du liquide blastocoelique et
de la multiplication cellulaire, le blastocyste commence d'augmenter en taille, entraînant
d'abords l'amincissement de la zone pellucide puis sa rupture.
La masse cellulaire interne est ensuite le siège de remaniements morphologiques et
d'une différenciation cellulaire menant à la mise en place des trois feuillets primitifs :
l'ectoderme, le mésoderme et l'endoderme. Cette phase est dénommée gastrulation. La
fonction de ces trois feuillets peut grossièrement se schématiser comme suit :
- Ectoderme : protection et réception des stimuli
- Endoderme : digestion et respiration
- Mésoderme : support, mouvement, excrétion et reproduction

3.2 Clivage et gastrulation chez les oiseaux
3.2.1 Le clivage
L'œuf macrolécithe de l'oiseau présente une segmentation méroblastique et
télolécithe. Les abondantes réserves de vitellus seront utilisées durant l'incubation et les
premiers jours de la vie aérienne (cette caractéristique permet la commercialisation des
poussins "day one" sans avoir besoin de les nourrir ni de les abreuver).
La fécondation de l'ovule n'est possible que durant les 15 minutes qui suivent
l'ovulation. A l'issue de la syngamie, le noyau zygotique se trouve en périphérie de l'énorme
cellule contenant le vitellus. Il est entouré d'une couche de vitellus blanc, au sommet de la
latébra. Trois à cinq heures après la fécondation, un premier sillon de clivage apparaît à la
surface du futur blastodisque, suivi 20 minutes plus tard d'un second sillon disposé à angle
droit (figure 3.3). Si la caryocinèse est complète, la cytocinèse reste inachevée, donnant au
blastodisque en formation l'aspect d'un syncitium. Des sillons radiaux se mettent en place,
isolant progressivement les premiers blastomères qui maintiennent cependant un contact avec
le vitellus sous-jacent. Un plan de clivage horizontal finit par les isoler complètement (figure
3.4). Le blastodisque est alors constitué d'une zone centrale de blastomères indépendants de
petite taille, le blastoderme, lui-même entouré de blastomères plus grands et incomplètement
individualisés. Les mitoses des blastomères centraux aboutissent à la mise en place de
plusieurs assises cellulaires et à l'apparition d'une cavité sous germinale par soulèvement du
blastoderme. L'embryon est alors qualifié de blastula primaire. Ainsi, la croissance du
blastoderme se fait par division des cellules centrales et par addition de blastomères
marginaux. Les bords du blastodisque conservent un aspect hyalin du fait de leur nature
syncitiale.
Au sein du blastoderme, les blastomères centraux se séparent en deux sous-
populations de cellules de taille différente, initialement mélangées. Les grosses cellules
riches en vitellus s'isolent des plus petites pour former le toit de la cavité sous-germinale et
s'accumulent préférentiellement à un endroit du blastodisque (figure 3.4). Cet endroit marque
la future extrémité caudale de l'embryon. De ce massif, des cellules migrent pour recouvrir le
plancher de la cavité sous-germinale. Elles sont rejointes par des cellules qui se détachent du
plafond. L'ensemble finit par former une assise cellulaire recouvrant complètement le
plancher de la cavité : l'hypoblaste. Les cellules du plafond de la cavité constituent
l'épiblaste. La cavité est maintenant complètement délimitée par des cellules et prend le nom
de cavité blastocoelique. L'embryon est arrivé au stade de blastula secondaire (figure 3.4).

Vu du dessus, la partie centrale du blastoderme a un aspect transparent de par la
présence de la cavité blastocoelique et prend le nom d'aire pellucide, tandis que la partie
périphérique est appelée aire opaque. L'embryon se développe uniquement à partir du disque
embryonnaire constitué de la partie centrale du blastoderme. Le reste du blastoderme formera
les organes extraembryonnaires.
3.2.2. Etablissement du plan de symétrie bilatérale
La formation du massif de cellules riches en vitellus se fait suivant le sens de rotation
de l'oeuf au cours de son trajet dans le segment inférieur de l'oviducte de la poule (ou la
cane). Cette rotation, responsable de la torsion des chalazes, se fait toujours dans le même
sens. Il en résulte que, si l’œuf progresse le petit bout en avant (90% des cas), l'orientation de
l'embryon est dite conforme à la loi de Von Baer, c’est-à-dire qu'en plaçant l’œuf
horizontalement, le blastodisque vers le haut et le petit bout à droite; l'axe longitudinal de
l'embryon est perpendiculaire au grand axe de la coquille et le pôle céphalique opposé à
l'observateur. Si l'oeuf transite le gros bout en avant (10 % des cas), la tête de l'embryon se
trouve face à l'observateur.
La définition de ce premier plan de symétrie crânio-caudale et la position relative de
l'épiblaste (en surface) et de l'hypoblaste (en profondeur) déterminent les deux autres plans
(dorso-ventral et bilatéral (gauche - droite)). Vingt-quatre heures après la fécondation, les
trois axes fondamentaux sont définis.
3.2.3. Chronologie de l'embryogenèse du poulet.
La période de segmentation couvre les 24 heures séparant la fécondation de
l'ovoposition (ou ponte). La gastrulation ne débute qu'après le ponte.
3.2.4. La gastrulation
3.2.4.1. Apparition de la ligne primitive et du nœud de Hensen
Le disque embryonnaire s'allonge suivant l'axe longitudinal. Les blastomères de
l'épiblaste convergent vers la région médio-caudale de l'aire pellucide pour y former un
bourrelet médian, premier stade de la ligne primitive. Cette ligne primitive s'allonge vers le
pôle craniâl au fur et à mesure de l'accumulation des cellules à sa base. Elle marque
l'emplacement du futur névraxe (SNC et moelle épinière). L'extrémité craniâle de cette ligne
est légèrement renflée et porte le nom de nœud primitif ou nœud de Hensen (figure 3.5).

3.2.4.2. Formation de l'endoderme et du mésoderme
Au centre de la ligne primitive se creuse un sillon par invagination des cellules
épiblatiques : la gouttière primitive. Les cellules de l'épiblaste qui s'invaginent se
différencient en cellules mésodermiques et rejoignent la cavité blastocoelique pour constituer
une assise cellulaire séparant l'épiblaste de l'hypoblaste. Cette nouvelle assise constitue le
mésoderme. Ce feuillet mésodermique s'étale latéralement et antérieurement pour entourer
l'extrémité craniâle de la ligne primitive (figures 3.5 et 3.6). La cavité blastocoelique
disparaît progressivement.
Les cellules mésodermiques qui rejoignent l'hypoblaste le long de la ligne primitive
s'insinuent entre les cellules hypoblastiques et les déplacent vers la périphérie de la cavité
blastocoelique. Elles se différencient en endoderme embryonnaire. Les cellules
hypoblastiques refoulées deviennent l'endoderme extraembryonnaire (figure 3.7).
3.2.4.3. Régression de la ligne primitive et formation de la notochorde
La ligne primitive cesse de s'allonger par son extrémité craniâle car les cellules
épiblastiques qui s'invaginent pour former le mésoderme n'y sont plus remplacées. Le disque
embryonnaire, lui, continue de s'étendre si bien que la ligne primitive paraît régresser et le
nœud de Hensen recule.
Au cours de ce mouvement, les cellules mésodermiques du nœud de Hensen élaborent,
sous l'épiblaste, un axe mésodermique médian qui s'étire par le recul du nœud et l'allongement
du disque embryonnaire: la notochorde (figure 3.6).
La gastrulation aviaire commence quelques heures après l'ovoposition, soit 24 à 30
heures après la ponte ovulaire. La ligne primitive atteint sa longueur maximale (2 mm) 18
heures plus tard et disparaît après 2,5 jours d'incubation.
Lorsque la ligne primitive a complètement régressé et qu'aucune cellule
mésodermique n'est plus produite par l'épiblaste, la gastrulation est terminée. Les cellules
épiblastiques restées en surface forment l'ectoderme.
Les marges latérales du mésoderme constituent le mésoderme latéral. Elles s'étendent
latéralement et rostralement pour se rejoindre sur la ligne médiane en avant de l'embryon. Ce
mésoderme latéral se sépare en deux feuillets : le feuillet pariétal ou somatique et le feuillet
viscéral ou splanchnique. Ces deux feuillets délimitent deux nouvelles cavités qui
fusionneront par la suite pour constituer le coelome. Le mésoderme latéral participe à la
formation des annexes extraembryonnaires ainsi qu'au système musculaire et cutané du
fœtus. Les dernières cellules mésodermiques produites demeurent sous l'ectoderme, près de
l'axe médian et forment le mésoderme paraxial qui est à l'origine des somites (musculature,
squelette, derme). Chaque somite sera relié au mésoderme latéral par un étroit pont
mésodermique : le néphrotome. Ce dernier est à l'origine des reins et des gonades.

3.3. Le clivage et la gastrulation chez les mammifères
3.3.1. le clivage
L'ovule de l'embryon de mammifère euthérien est alécithe (ou oligolécithe) et la segmentation
de l'embryon est holoblastique et isolécithe. L'entièreté du cytoplasme est divisée et les
blastomères sont tous de taille comparable (figure 3.1).
Le clivage se déroule dans l'oviducte (ou trompe de Fallope) et la taille des
blastomères diminue au fur et à mesure des mitoses successives puisque le volume total de
l'embryon ne se modifie pas. Les premiers clivages sont sous la dépendance des ARNm
maternels stockés durant la maturation de l'ovocyte. Ce n'est qu'au stade 8-16 cellules chez
les bovins que le génome embryonnaire prend le contrôle du développement. Cette transition
est associée à un arrêt momentané des divisions, à la disparition des transcrits maternels et à
l'apparition des transcrits embryonnaires. Le’”réveil" du génome embryonnaire se produit au
stade 2 cellules chez la souris, 4-8 cellules chez le lapin et 8-16 cellules dans l'espèce
humaine.
Les caractéristiques générales du cycle cellulaire(1) s'appliquent au clivage
embryonnaire, mais la durée des différentes phases change au cours des divisions
successives. Chez les bovins et les ovins, le premier cycle est long, les suivants sont courts
jusqu'au moment de la transition du contrôle maternel au contrôle embryonnaire, après quoi
les cycles redeviennent courts. Ce sont surtout les phases G1 et G2 dont les durées sont
modulées.
(1) Le cycle cellulaire de toutes les cellules en croissance et en division a une durée de 10 à 20 heures et
comprend deux étapes (figure 3.8) :
a) la division et la séparation des cellules filles (phase M et cytocinèse)
b) l'interphase ou période de croissance divisée elle même en périodes G0 et G1, phase S et période G2.
La période G0, quand elle existe, correspond à un état de quiescence mitotique. La cellule cesse de se diviser
(ex. les neurones, les fibres musculaires striées). La période G1 correspond à la croissance cellulaire
(accroissement des réserves, synthèse de macromolécules, multiplication des organites, division des
mitochondries, augmentation de la taille cellulaire). La phase S (pour synthèse) se caractérise par la duplication
de l'ADN et la synthèse des histones et d'autres protéines liées à l'ADN. La croissance cellulaire se poursuit. La
phase G2 prépare la cellule à l'accomplissement de la mitose proprement dite.
Le cycle cellulaire est un des grands sujets de recherche de la biologie fondamentale car la connaissance fine de
son contrôle moléculaire mènera à mieux comprendre d'autres processus tels que celui de l'équilibre
différenciation-division, celui de la cicatrisation ou encore celui de la "cancérisation".
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%