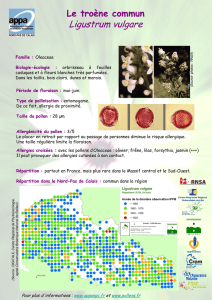
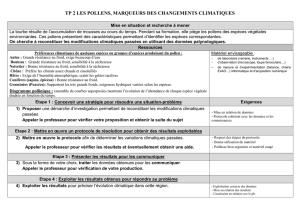
QUATORZIEME SEMINAIRE BOTANIQUE ET ALLERGIE

QUATORZIEME SEMINAIRE BOTANIQUE ET ALLERGIE DE TOULON
FLORE COTIERE ET INSULAIRE MEDITERRANEENNE
Isabelle Sullerot pour le texte, Laurent Bray pour les photos de plantes et Jean Paul Masy pour les
photos d’ambiance
LaquatorzièmeéditiondecetévénementincontournableaeulieuàToulon,du8au10mai
2015.Jean‐FrançoisFontaineetlePrFrédericDupontonteneffetreprisleflambeauetrelevéle
défi,avecleconcoursdeslaboratoiresALKtoujoursfidèles,etceluidulaboratoireMEDAquia
acceptéderejoindrel’aventure.Etl’aventurefutmagnifique,servieparunemétéoestivalequi
adonnétoutesavaleuràcetterégionsplendide,dontlejoyau,l’îledePortCrosfutlepoint
d’orgue.
Lesparticipantssesontretrouvéslevendredisoir,certainsvenantsdetrèsloin,autourd’un
dînerconvivialaucoursduquelleprogrammedesdeuxjournéesaétédétaillé.Lesamedimatin
étaitconsacréauxconférencesscientifiques,etdeuxsortiesbotaniquesétaientorganiséesle
samediaprès‐midietledimanchematin.
Enouverturedessessionsscientifiques,Jean‐FrançoisFontaineetJean‐MarcDevoisinsont
expliquéleurattachementàceséminaireoriginal,réunissantallergologues,botanisteset
universitaires,autourdepromenadesbotaniques,véritablestravauxpratiques«insitu»,grâce
àlavolontédestroisélémentsmoteursquesontl’ANAFORCAL,LesJardinsBotaniquesde
France,etlespartenairesdel’industriepharmaceutique.
COMPTE- RENDU DES CONFERENCES
Du pollen à la pollinose : Pr Gabrielle Pauli
Pourquoicertainspollenssont‐ilsplusallergisantsqued’autres?LepostulatdeThommen
donnetroisréponses:lepollendoitcontenirunprincipeantigénique,êtreanémophile,
abondantetapparteniràuneplantesuffisammentcommunedanslarégionconsidérée.
Lespollensanémophilessontaérodynamiques,émisengrandesquantités,etsurdelongues
distances.Ilsappartiennentàdesplantesvivantenpopulationsserrées,dontlesfleurssontnon
colorées,sansodeur,ninectar.
Lepotentielallergisantd’unpollenestfondésurlaquantitéd’allergènesetlafréquencedes
sensibilisationsdansunerégiondonnée:enAlsacelasensibilisationauxgraminéesest
supérieureàcelledesbouleaux,cequidémontreunpotentielintrinsèquesensibilisant
supérieurpourlesgraminées.
Lerisqueallergiqueestunedonnéesanitairequiestlarésultantedesdifférentsparamètres:
potentielallergisantdupollen,zonegéographiqued’émission,quantitédepollensdansl’air,
conditionsmétéorologiques,observationscliniques.Parexemple,lepollendebouleaudontle
potentielallergisantestde5,aunrisqueallergiquede5/5àStrasbourg,maisde1/5àNice.Et

inversementpourlepollendecyprès.DanslafamilledesCupressacées,lecyprèsetlegenévrier
ontunrisqueallergiquedemoyenàélevé,tandisquelethuyaaunrisquefaible.Danslafamille
desPinacées,lesmélèzes,pins,sapins,cèdres,ontunrisqueallergiquefaiblebienqu’ilssoient
majoritairessurlescomptespolliniques.
Lerisqueallergiquepeutêtremodifiédansletemps:pourlecyprèsilatteintentre2,4et8%,
alorsqu’en1970,onpubliait3casd’allergieaucyprès(Aubert,Marseillemédical).AStrasbourg,
4%desenfants(10à12ans)sontsensibilisésaubouleau,tandisqu’en1976,unepublication
faisaitétatde31casen5ans(Pauli,Bessot,Bulletind’Actualitésthérapeutiques).
Maisilexistedespollinosesdeproximité,d’espècespeuévocatrices(chèvrefeuille),etilfaut
réunir5cascliniquesargumentéspourretenirlerôleallergisantd’unpollen.
L’allergénicitédupollendépenddenombreuxfacteursconditionnantl’abondanceauniveau
respiratoire:desfacteursphysiques(nombreettailledesgrains,surfacelisseoucollante,
flottaisondansl’air),etdesfacteursimmunochimiques(tauxvariabledeprotéinesallergéniques
ounon).
Leprincipalfacteurphysiquedéterminantl’allergénicitéestlasolubilité,ainsiqu’unPMentre
10et100KDasuffisantpourstimulerlescellulesimmunocompétentes,etpastropgrandpour
lepassageàtraverslesmuqueuses.Parexemple,l’allergèneBetv1(allergènemajeur),estpeu
abondantdansl’airinhalé,maistrèsprésentdanslesurnageantdepollendebouleauhydraté,
dèsla10émeminute,etenconcentrationcroissantejusqu’àla6èmeheure.Contrairementàla
glutathionStransférase,(allergènemineur:13%desensibilisations)abondantedanslepollen
debouleau,maispeulibéréedanslamuqueusenasalecarpeuhydrosoluble.
Lesautresdéterminantssontlafonctionenzymatique,lacapacitédetranslocationaucontact
desmembranescellulaires(quiestliéeàuntransfertactifdelapartdescellulesépithélialesdes
sujetsallergiques),etl’activitéTh2oupro‐inflammatoire.Celle‐ciestliéeàlafonctionde
l’allergène:
- lesenzymes(pectasesliases,isoflavone‐réductases,ribonucléases,chitinases,...)favorisentla
pénétrationdesprotéinesallergéniques,etinteragissentaveclesrécepteursPARprésentssur
lescellulesépithélialesquisontalorsactivées.
‐LaprésencedePALMS(pollenassociatedlipidmédiators)dansl’exineetl’intine:acidesgras,
n—alkanesquistimulentlesréponsesimmunes,etamplifientlaréponseauxprotéines
allergéniques.
Enfin,l’allergénicitédupollenpeutêtremodifiéeparl’environnementextérieur:lerôlede
l’ozoneaétédémontrépourlepollendebouleau,etceluiduC02pourlepollend’ambroisie.
De la pollinose à l’allergie alimentaire. Dr Jean François Fontaine
Lesallergiescroiséespollens–alimentssontfréquentes;leursmanifestationscliniquessontde
gravitédiverse,allantdusimplesyndromeoralauchocanaphylactique;ellessontduesàdes

famillesd’allergènescommunesaurègnevégétal.
Ondistinguelespanallergènes, ubiquitaires,leseurallergènes, delargedistribution,les
stenallergènes, dedistributionpluslimitéeetderéactionscroiséesmoinsconstantes,etles
monallergènes limitésàunesourceallergénique(ex:lespolcalcinesuniquementprésentes
danslespollens).
1.Lesprofilines
Panallergènes,protéinesducytosquelettedescelluleseucaryotes,présentesdansdenombreux
pollensetaliments,dontl’homologiedeséquenceestgrandeentrelesespèces,elles
sensibilisent20%despolliniques,etsontresponsablesdemultiplesréactionscroiséesentre

pollensd’espèceséloignées,etentrepollensetaliments.
Tableau1
2.LesprotéineshomologuesdeBetv1
ProtéinesdedéfensedesvégétauxPR10,eurallergènespolliniquesetalimentaires,dont
l’homologiedeséquenceestgrandeentrelesespèces,présentesdanslespollensdebétulacées,
decorylacées,defagacées,etdanslesvégétauxcomestibles:pomme,pêche,cerise,noisette,
carotte,céleri,arachide,soja,kiwi,fruitdujacquier,...,sanstraductioncliniquesystématique.
3.Lesprotéinesdetransfertlipidique(LTP)
ProtéinesdedéfensedesvégétauxPR14,thermostablesetrésistantesàlaprotéolyse,
eurallergènesprésentsdanstouteslescellulesvégétales(pollens,tiges,fleurs,feuilles,fruits),
enplusgrandequantitédanslapeaudesfruitsquedanslapulpe,onlesretrouvedanslesjusde
fruits,lesvins,lesconfitures,danslescéréales,leslégumineusesetlesfruitsàcoque,etlelatex,
lecannabis,...dontl’homologiedeséquenceestfaibleentrelesespèces.
Encequiconcernelespollens,leursensibilisationestvariable,etleurimplicationcliniqueest
discutée..Lespollensdeplataneetd’armoisesontplussouventconcernésqueceuxdecyprès.

Encequiconcernelesaliments,ellessontresponsablesdemanifestationscliniquesparfois
sévères.Danslesyndromepêche‐pollen,lasensibilisationLTPdel’armoiseestconsécutiveà
celledelaLTPdelapêche,etlapollinoseàl’armoiseresteasymptomatique;inversementen
Chineoùlasensibilisationprimaireestl’armoisedanslesrégionsoùcepollenesttrèsabondant.
4.Lesthaumatines‐likeprotéines(TLP)
ProtéinesdedéfensedesvégétauxPR5,thermostablesetrésistantesàlaprotéolyse,présentes
dansdiverspollens(cupressacées,platane)etvégétauxcomestibles(pêche,kiwi,raisin,banane,
brocoli),stenallergènes,supportderéactionscroiséesplusinconstantes.
5.1‐3ßglucanases
ProtéinesdedéfensePR2,présentesdansdiversvégétaux,pollens,latex,ellessont
responsablesderéactionscroiséespollens‐aliments,maissanstraductioncliniqueidentifiée.
6.Lesisoflavonesréductases
Présentesdansdenombreuxvégétaux,avecunegrandehomologiedeséquence,leur
pertinencecliniqueresteàdéterminer.
Conclusion:Lesassociationspollinosesetallergiesalimentairessontfréquentes,certaines
réactionscroiséessontidentifiées,d’autresrestentàdécouvrir.Lasensibilisationpolliniqueest
primairedanscertainscas(<PR10),avecunrôleprobabledela«pressionpollinique»;elle
peut‐êtreunmarqueurdesensibilisationmoléculairedansd’autrescas(LTP).
Entomophagie et risque allergique. Drs Etienne Beaudouin et François Lavaud
Laconsommationd’insectesparl’êtrehumainestunepratiquetrèsancienne,surtoutdansles
régionstropicalesdel’Afrique,l’Amériquelatineetl’Asie.Elledevientàlamodedanslespays
occidentaux,oùsedéveloppentdessitesdeventesurinternet,etdesrestaurantsdédiés.
Lesbénéficesavancésdel’entomophagiesontdenourrirlapopulationmondialeavecun
rendementmaximum,uneconsommationd’eauminimale,etuneproductiondegazàeffetde
serreréduite.
Lesavantagesnutritionnelssontunerichesseenprotéines,encertainsacidesaminés,en
vitamines,enlipides,avecunratioacidesgrassaturés/insaturésidéal,etunepauvretéen
glucides.
Lesinsecteslesplusconsomméssontlescoléoptères(charançondupalmier,verdefarine),les
lépidoptères(chenillemopane,chenilledubambou),leshyménoptères(fourmis,guêpes:larves
etnymphes),lesorthoptères(criquets,grillons,sauterelles),leshémiptères(cigales,citadelles,
cochenilles),lesisoptères(termites).
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%