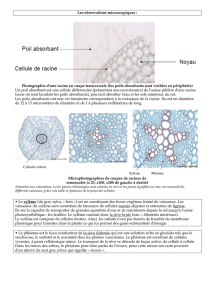
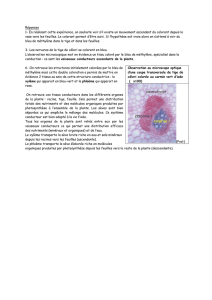
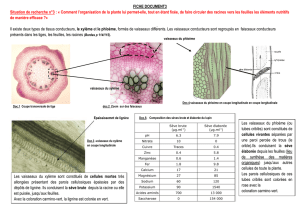
TRANSLOCATION DANS LE PHLOEME

Relations plante et eau A.Hafidi (a.haf[email protected])
UNIVERSITE CADI AYYAD
FACULTE DES SCIENCES SEMLALIA
DEPARTEMENT DE BIOLOGIE
RELATIONS PLANTE ET EAU

Relations plante et eau A.Hafidi (a.haf[email protected])
Chapitre 0 : PROPRIETES DE L’EAU ET NOTIONS DU
POTENTIEL HYDRIQUE
0.1 Structure et propriétés de l’eau
0.1.1 Les liaisons hydrogènes
0.1.2 Les propriétés thermiques
0.1.3 Les propriétés de cohésion et d'adhésion
0.1.4 La résistance à la distension
0.1.5 Le potentiel chimique de l'eau
0.2 L’eau dans le sol
0.2.1 Le potentiel hydrique du sol
0.2.2 Le flux (écoulement) de masse
0.2.3 La conductivité hydraulique du sol
0.2.4 Le point de flétrissement permanent
0.3 L’eau dans la plante
0.3.1 La pression de turgescence
0.3.2 Le potentiel hydrique au sein de la plante
0.3.3 L’équilibre hydrique de la plantes

Relations plante et eau A.Hafidi (a.haf[email protected])
De toutes les ressources dont les plantes ont besoin pour se développer, l'eau est la plus abondante et en
même temps la plus limitante pour les productivités agricoles. L'eau constitue en général l'essentiel (70 à 95%)
de la masse des tissus végétaux. En fait, pour une photosynthèse, active, elles ont besoin d’ouvrir leurs stomates
pour absorber le dioxyde de carbone de l'atmosphère, ceci les expose à la perte d'eau et la menace de
déshydratation. Les plantes doivent ainsi en permanence absorber et perdre de l'eau. La transpiration sert à
dissiper la chaleur générée par les rayons du soleil et l’activité catabolique. En outre, le flux d'eau absorbée par
les racines permet de transporter les minéraux dissouts.
0.1 Structure et propriétés de l’eau
L'eau a des propriétés particulières qui lui permettent d'agir comme un excellent solvant et d’être
facilement transportée à travers la plante. Ces propriétés sont dues essentiellement à la structure polaire de la
molécule d'eau (Fig. 1). Les charges partielles opposées entre molécules d'eau voisines ont tendance à s’attirer.
Ces liaisons électrostatiques faibles entre les molécules d'eau sont dites liaisons hydrogènes et sont à l’origine de
propriétés importantes de l’eau.
Fig.1 : Schéma d’une molécule d’eau
Fig. 2 : liaisons hydrogènes entre molécules d’eau
0.1.1 Les liaisons hydrogènes
La polarité de l'eau en fait un excellent solvant, elle dissout de grandes quantités d'une grande variété
de substances. Les liaisons hydrogènes (Fig. 2) entre les molécules d'eau et les ions, et entre l'eau et les solutés
polaires, en solution diminuent l'interaction électrostatique entre les substances chargées et donc augmentent
leur solubilité.
0.1.2 Les propriétés thermiques
Chaleur spécifique élevée : Lorsque la température de l'eau est élevée, les molécules vibrent plus
rapidement et avec une plus grande amplitude. Pour permettre ce mouvement, l'énergie ajoutée au système doit
briser les liaisons hydrogène entre molécules d'eau. Ainsi, comparativement à d'autres liquides, l'eau nécessite
relativement un apport d'énergie plus grand pour élever sa température.
Chaleur latente de vaporisation élevée : A 25 ° C, elle est de 44 kJ mol-1. C’est la plus haute valeur
connue pour les liquides. La plupart de cette énergie est utilisée pour casser les liaisons hydrogène entre
molécules d'eau. La forte chaleur latente de vaporisation d'eau permet aux plantes de se rafraîchir par
évaporation de l'eau à la surface des feuilles.
0.1.3 Les propriétés de cohésion et d'adhésion
Les molécules d'eau à l'interface air-eau sont plus fortement attirées par les molécules d'eau voisines
que par la phase gazeuse donnant naissance à une tension de surface. Les multiples liaisons hydrogène entre les
molécules de l'eau donnent également naissance à la propriété connue sous le nom de Cohésion. Il s’agit de
l'attraction mutuelle entre les molécules d’eau. Une propriété analogue, appelée Adhérence, est due à
l'attraction des molécules l'eau par une surface solide comme les parois des cellules ou une surface en verre. La
tension de surface, les forces de cohésion et d’adhésion sont responsables du phénomène de capillarité.
Chapitre 0 : PROPRIETES DE L’EAU ET NOTIONS DU POTENTIEL HYDRIQUE
2

Relations plante et eau A.Hafidi (a.haf[email protected])
0.1.4 La résistance à la distension
Les forces de cohésion donnent à l'eau une résistance à la distension élevée, définie comme étant la
force maximale par unité de surface que peut supporter une colonne continue d’eau avant de casser. Briser la
colonne d'eau nécessite de l'énergie suffisante pour rompre les liaisons hydrogènes qui attirent les molécules
d'eau entres elles.
0.1.5 Le potentiel chimique de l'eau
Le potentiel chimique de l'eau est une expression quantitative de l'énergie libre associée à une mole de
molécules d'eau. Pour des raisons historiques, en physiologie végétale, un paramètre similaire appelé potentiel
hydrique, est souvent utilisé. Il est défini comme le potentiel chimique de l'eau divisé par le volume molaire
partiel de l'eau (le volume d'une mole de l'eau): 18 × 10-6 m3 mol-1. Les principaux facteurs influençant le
potentiel hydrique des plantes sont la concentration, la pression hydrostatique et la gravité. Le potentiel
hydrique des solutions est symbolisé par Ψw, et peut être subdivisé en composantes individuelles : Ψw
=Ψs+Ψp+Ψg . Les termes Ψs, Ψp, et Ψg désignent les effets des solutés, de la pression hydrostatique, et de la
gravité, respectivement. Le terme Ψs, appelée potentiel soluté ou potentiel osmotique, représente l'effet des
solutés dissouts sur le potentiel hydrique. Le potentiel osmotique est indépendant des spécificités et de la nature
du soluté.
Le terme ΨP est la pression hydrostatique de la solution. Les pressions positives font monter le potentiel
hydrique, les pressions négatives le réduisent. La pression hydrostatique positive dans les cellules est la pression
appelée pression de turgescence. La gravité fait tirer l’eau vers le bas. Le terme Ψg dépend de la hauteur (h) de
l'eau, de la densité de l'eau (ρw), et l'accélération de la pesanteur (g). Ψg = ρw gh où ρwg a une valeur de 0,01
MPa m-1. Ainsi, une distance verticale de 10 m se traduit par un changement de 0,1 MPa pour le potentiel
hydrique. Au niveau cellulaire, la composante gravitationnelle (Ψg) est généralement négligée par rapport au
potentiel osmotique et la pression hydrostatique. Ainsi, dans ce cas : Ψw =Ψs +Ψp
0.2 L’eau dans le sol
La teneur en eau et le flux de l'eau dans les sols dépendent dans une large mesure du type de sol et sa
structure. Quand un sol est fortement hydraté, l'eau percole vers le bas par gravité à travers les espaces entre les
particules du sol, déplaçant en partie, et en piégeant dans certains des cas, de l'air dans des canaux. L'eau dans
le sol peut exister sous forme d’un film adhérant à la surface des particules du sol, ou remplissant la totalité des
espaces entre les particules.
Dans les sols sableux, les espaces entre les particules sont si grands que l'eau a tendance à drainer
facilement, seule une fine couche peut rester sur les surfaces des particules et dans les interstices entre
les particules.
Dans les sols argileux, les interstices sont suffisamment petits pour que l'eau ne puisse circuler
librement entre eux, elle est tenue plus étroitement par des forces de capillarité et d’adhésion.
La capacité de rétention de l’eau des sols est appelée la capacité au champ. La capacité au champ est la teneur
en eau d'un sol après qu'il ait été saturé d'eau et que l'excès d'eau ait été laissé drainer. Les sols argileux ou les
sols avec une haute teneur en humus ont une capacité au champ élevée. En revanche, les sols sableux
généralement ne retiennent que peu d’eau.
0.2.1 Potentiel hydrique du sol
Comme le potentiel hydrique des cellules végétales, le potentiel hydrique des sols peut être subdivisé en
deux composantes : le potentiel osmotique et la pression (potentiel) hydrostatique. Le potentiel osmotique (Ψs)
de l'eau dans le sol est généralement négligeable parce que les concentrations de solutés sont faibles. Une valeur
typique pourrait être -0,02 MPa. Cependant, pour les sols qui contiennent une importante concentration de
sels, Ψs est significatif, et peut-être de -0,2 MPa ou encore plus faible. La deuxième composante du potentiel
Chapitre 0 : PROPRIETES DE L’EAU ET NOTIONS DU POTENTIEL HYDRIQUE
3

Relations plante et eau A.Hafidi (a.haf[email protected])
hydrique du sol est la pression hydrostatique (Ψp). Pour les sols humides, Ψp est très proche de zéro. Quand un
sol se dessèche, Ψp diminue et peut devenir assez négatif.
0.2.2 Le flux (écoulement) de masse
L'eau se déplace à travers les sols principalement par le flux de masse entraîné par un gradient de
pression. Quand les plantes absorbent l'eau du sol, elles épuisent les sols de l'eau près des racines. Cet
appauvrissement réduit le ΨP de l'eau près des racines et établit un gradient de pression avec régions voisines
du sol qui ont des valeurs supérieures Ψp. Les espaces remplis d'eau sont interconnectés, l'eau se déplace alors
jusqu’à la surface de la racine. La vitesse et le taux d'écoulement d'eau dans les sols dépend de deux facteurs:
l’intensité du gradient de pression à travers le sol, et la conductivité hydraulique du sol.
0.2.3 La conductivité hydraulique du sol
La conductivité hydraulique du sol étant une mesure de la facilité avec laquelle l'eau se déplace à
travers le sol, elle varie avec le type de sol et sa teneur en eau. Les sols sableux, avec leurs grands espaces entre
les particules, ont une conductivité hydraulique grande, tandis que les sols argileux, ont une conductivité
hydraulique nettement plus petite.
0.2.4 Le point de flétrissement permanent
Dans les sols très secs, le potentiel hydrique (Ψw) peut tomber en dessous de ce qu'on appelle le point de
flétrissement permanent. A ce niveau le potentiel hydrique du sol est si faible que les plantes ne peuvent pas
retrouver la pression de turgescence, même si toutes les pertes d'eau par la transpiration cessent.
0.3 L’eau dans la plante
L'eau pénètre ou sort de la cellule selon le gradient de potentiel hydrique. Le transfert de l'eau est un
processus passif. L'eau se déplace en réponse à des forces physiques, vers les régions à faible potentiel hydrique
ou à faible énergie libre. Toutefois, Dans le phloème, l'eau peut être transportée de régions à faibles potentiels
d'eau (par exemple, les feuilles) vers des régions où le potentiel hydrique est plus élevé (par exemple, les racines).
0.3.1 La pression de turgescence
Les cellules végétales ont des parois assez rigides, tout changement du potentiel hydrique Ψw des
cellules est généralement accompagné par un grand changement du Ψp, avec relativement peu de changements
du volume des cellules (Protoplastes). Ce phénomène est illustré dans les courbes de Ψw, Ψp, et Ψs en fonction
du volume relatif de la cellule (Fig. 3).
0.3.2 Le potentiel hydrique de la plante
Tout comme le Ψw, le Ψs peut varier considérablement en fonction des conditions de croissance et le
type de plante. Dans les cellules de plantes bien hydratées, Ψs peut atteindre -0,5 MPa, mais généralement des
valeurs de -0,8 à-1,2 MPa sont plus normales. Les plantes en conditions de sécheresse peuvent parfois atteindre
des Ψs très bas. Le déficit hydrique conduit généralement à une accumulation de solutés dans le cytoplasme et
la vacuole, permettant ainsi de maintenir la pression de turgescence de la plante, malgré des potentiels
hydriques très bas. Les tissus végétaux satockant des concentrations élevées de saccharose ou d'autres sucres,
tels que les racines de betterave à sucre, les tiges de canne à sucre, ou les baies de raisin, atteignent des valeurs
très basses de Ψs de l’ordre de -2,5 MPa. Les plantes halophytes, montrent habituellement des valeurs très
faibles de Ψs. Un Ψs faible abaisse le Ψw cellulaire suffisamment pour extraire de l'eau pure de l'eau salée, sans
permettre à des niveaux excessifs de sels d'entrer en même temps dans la cellule.
Une pression de turgescence positive (Ψp) est importante pour deux raisons principales. Premièrement, la
croissance des cellules végétales nécessite une pression turgescence pour étirer les parois cellulaires. La deuxième
Chapitre 0 : PROPRIETES DE L’EAU ET NOTIONS DU POTENTIEL HYDRIQUE
4
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
1
/
27
100%