Role du mesoderme somitique dans le developpement du plumage

/.
Embryol.
exp. Morph. Vol. 28, 2, pp. 313-341,1972 313
Printed in Great Britain
Role du mesoderme somitique
dans le developpement du plumage
dorsal chez Tembryon de Poulet
I. Origine, capacity's de regulation et determination
du me'soderme plumigene
Par ANNICK MAUGER1
Laboratoire de Zoologie de VUniversite scientifique
et medicate de Grenoble
SUMMARY
The role of
somitic
mesoderm in the development of
dorsal
plumage in chick
embryos. I. Origin, regulation capacity and determination
The role of somitic mesoderm in the development of the dorsal plumage has been studied
in chick embryos. The operations were performed at 2-2-5 days of incubation.
The replacement of a portion of somitic mesoderm by somitic mesoderm labelled with
[3H]thymidine or obtained from Japanese quail embryos (whose nuclei bear distinctive
specific markers) showed that cells originating from the dermatomes build up the dermis of
the dorsal skin only. They do not migrate farther than approximately midway down the
flank. Beyond this limit, dermal cells originate from the somatopleural mesoderm.
The unsegmented somitic mesoderm is capable of extensive regulation, which leads to the
development of a dorsal plumage, normal in the number and arrangement of its feathers
according to the characteristic pattern of the spinal pteryla. Uni- or bilateral excision of
segmented somitic mesoderm resulted in dorsal plumage deficiencies, the extent and fre-
quency of which was related to the state of differentiation of the excised mesoderm. Thus,
the excision of somites generally led to an incomplete spinal pteryla (absence of feather rows,
apteria). However, the somitic mesoderm is still capable of regulation even though it has
already undergone its differentiation into dermatome, myotome and
sclerotome.
These results
show that somitic mesoderm retains its regulative capacity, even though it has already ac-
quired its feather-forming determination.
The replacement of unsegmented somitic mesoderm by various implants (agar, tantalum,
gut, neural tube, somatopleural mesoderm), intended to block the regulation processes,
abolished the differentiation of the spinal feathers on the operated side. In some cases, the
implantation of somatopleural mesoderm resulted in the formation of a supernumerary tract.
No tissue other than somitic mesoderm - not even the somatopleural mesoderm, which is
normally in part feather-forming - is able to give rise to region-specific spinal pteryla dermis.
The excision and replacement of somitic mesoderm prevented the differentiation of dense
dermis, whereas these operations had no effect on the early histogenesis of the epidermis,
with the formation of arches and anchor filaments.
1
Adresse
deVauteur:
Laboratoire de Zoologie, Universite scientifique et medicale de Gre-
noble, 38-Saint-Martin-d'Heres, France.

314 A. MAUGER
INTRODUCTION
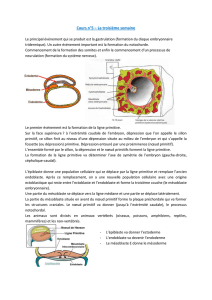
Le territoire presomptif de la pteryle spinale de l'embryon de Poulet de 2
jours d'incubation est entierement compris a l'interieur des bords externes des
somites (Mauger & Sengel, 1970). A l'interieur de ce territoire, l'ectoderme
donne naissance a l'epiderme et le mesoderme somitique dorsal (dermatome)
au derme de la peau dorsale (Rawles, 1955). A partir de 3 jours d'incubation,
le dermatome se desagrege et ses cellules remplissent Fespace sous-ectodermique
pour constituer le mesenchyme sous-ectodermique. Puis, a l'interieur des limites
de la future pteryle, les cellules predermiques se condensent uniformement sous
'lepiderme. Plus tard (entre 6,5 et 7,5 jours d'incubation) apparaissent les ebau-
ches plumaires de la pteryle spinale, dont la differenciation est le resultat d'inter-
actions inductrices entre le derme et l'epiderme (Sengel, 1958; Rawles, 1963).
Le derme et l'epiderme de la peau ventrale ou des membres sont egalement aptes
a edifier des pteryles (pectorale, ventrale, alaire ou femorale). Les cellules dermi-
ques de ces pteryles sont vraisemblablement issues du mesoderme somato-
pleural. En effet, des troncons de lames laterales, preleves au niveau des somites
20 a 25 (Straus & Rawles, 1953) ou du cordon somitique (Murray, 1928) et
greffees dans la cavite ccelomique ou sur la membrane chorioallantoidienne,
ont donne naissance a des plumes. Ainsi, le mesoderme somatopleural des
lames laterales est capable d'edifier un derme plumigene meme lorsqu'il est
explante avant l'emigration des cellules du dermatome. De plus, l'irradiation
des somites aux rayons X n'a jamais provoque de lesions des pteryles pectorale,
ventrale, alaire ou femorale (Mauger, 1970).
Bien que ces constatations permettent de penser que les cellules dermiques de
ces pteryles sont issues d'ebauches situees a Pexterieur des somites, une confirma-
tion experimental etait necessaire. Pour cela, des greffes de mesoderme somi-
tique marque a la thymidine tritiee ou des greffes de mesoderme somitique de
Caille (dont les noyaux se differencient aisement de ceux du Poulet (Le Douarin
& Barq, 1969)) ont ete entreprises afin de connaitre la destination des cellules
issues du dermatome.
L'irradiation aux rayons X d'une portion transversale du territoire presomp-
tif de la pteryle spinale (tube neural, somites adjacents et ectoderme sus-jacent)
empeche la differenciation des plumes dans le territoire cutane derivant de la
region irradiee. Des irradiations localisees sur le tube neural ou sur les somites
ont montre que la non-differenciation des plumes est liee a la radiolesion du
mesoderme somitique. Celui-ci, par suite de Felimination de nombreuses cellules
pycnotiques, montre, 24 heures apres l'irradiation, une forte diminution du
nombre des ses cellules. Toutefois, vers 5 jours d'incubation, le mesenchyme
sous-ectodermique a recouvre une densite cellulaire normale. Malgre cela il est
devenu incapable d'effectuer sa differenciation en derme dense (Mauger, 1970).
Diverses hypotheses ont ete avancees pour expliquer l'inaptitude du meso-
derme somitique irradie a former des plumes: les rayons X modifieraient le

Developpement du plumage dorsal. I 315
genome des cellules sous-ectodermiques, les rendant inaptes a se differencier en
derme dense; ou bien, les cellules mesodermiques rescapees de Pirradiation,
s'etant multipliees d'une facon anormalement rapide pour restaurer une densite
cellulaire normale, auraient perdu la capacite d'effectuer de nouvelles mitoses au
moment de P edification du derme dense.
Afin de verifier qu'une insuffisance numerique des cellules somitiques pouvait
etre a Porigine de la non-differentiation des plumes d'une portion de la pteryle
spinale, la deficience somitique a ete provoquee par un autre procede, qui
consiste a exciser chirurgicalement un troncon de mesoderme somitique. Ces
experiences ont montre que le mesoderme somitique est doue d'un grand pou-
voir de regulation, qui diminue cependant a mesure que Pintervention est
pratiquee a un stade plus tardif de la differenciation somitique. Puisque les
capacites de regulation du mesoderme somitique permettent la restauration
rapide de la deficience somitique provoquee par l'extirpation chirurgicale, le
materiel somitique excise a ete remplace par des implants d'origine variee,
destines a faire obstacle a la regulation au sein du mesoderme somitique.
De cette facon, le role du mesoderme somitique dans le developpement du
plumage dorsal a pu etre defini, en meme temps que ses capacites de regulation
et les modalites de sa determination ont ete elucidees.
MATERIEL ET METHODES
Les operations ont ete effectuees sur des embryons de Poulet de race Leghorn
blanche ages de 2 a 2,5 jours d'incubation (15 a 36 paires de somites; stades lla
18 de Hamburger & Hamilton (1951)).
(1) Techniques operatoires
(a) Extirpation chirurgicale du mesoderme somitique
Les embryons ont ete prealablement colores au rouge neutre (Hamburger,
1942).
Apres ressection de la membrane vitelline, les excisions ont ete prati-
quees a l'aide d'une aiguille courbe de verre noir. Elles ont ete effectuees a
divers niveaux de Paxe cephalo-caudal. Ainsi, le mesoderme somitique a ete
excise a differentes etapes de sa differenciation: au niveau du cordon somitique
insegmente sur une longueur de 6 a 9 somites presomptifs, immediatement en
arriere du dernier sillon intersomitique; au niveau des 5a 6 derniers somites;
au niveau des 5 a 6 somites situes en direction cephalique a une distance variant
de 3 a 12 somites en avant du dernier sillon intersomitique. Le mesoderme somi-
tique a ete excise de facon unilaterale (droite) ou bilaterale. Lorsque l'excision a
ete pratiquee au niveau du cordon somitique, celui-ci a generalement ete
elimine completement au-dessus du feuillet endodermique, qui reste intact
(Fig. 4). Au contraire, lorsque l'excision a ete pratiquee au niveau des somites, le
sclerotome a echappe, en partie ou completement, a l'extirpation (Fig. 5). II est,
en effet, difficile, sinon impossible, d'exciser completement les somites sans
provoquer d'hemorragie par rupture de l'une des aortes dorsales.
21 E M B 28

316A. MAUGER
Ectoderme
climine
Mesoderme
somatopleural
ventral
Ectoderme
' climine
Mesoderme
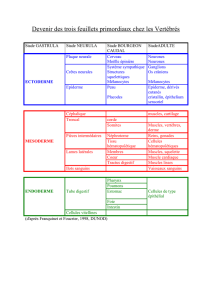
Fig. 1. Schema de l'operation qui consiste a remplacer le mesoderme somitique
par un autre tissu. A, implantation de mesoderme somatopleural distal (territoire
presomptif de l'apterie medio-ventrale). B, implantation de mesoderme somato-
pleural proximal (territoire presomptif de la pteryle pectorale). Le resultat de ces
operations est represents par les Figs. 18 (A) et 20 (B).
(b) Remplacement du mesoderme somitique
L'excision a generalement ete pratiquee aii niveau du cordon somitique droit.
Ce niveau a ete choisi en raison de la possibility d'extirper completement le
mesoderme somitique. Les embryons etaient ages de 12 a 28 paires de somites.
La plaie resultant de l'excision a ete immediatement comblee par un implant
de taille identique, soit inanime (tantale, agar), soit tissulaire vivant (tube
neural, tube
digestif,
mesoderme extra-somitique, mesoderme somitique marque,
mesoderme somitique de Caille) (Fig.
1
A, B).
Les implants inanimes etaient constitues par de petits parallelepipedes d'agar
melange a du noir animal, ou de tantale de 0,1 mm d'epaisseur.
Les implants de quartier de tube digestif ont ete preleves sur des embryons
de 4 a 6 jours d'incubation, disseques dans du liquide de Tyrode. Lors de la
greffe, F epithelium intestinal a ete place du cote de l'endoderme de l'hote. Les
autres greffons tissulaires vivants ont ete preleves sur des embryons de 2 a 2,5
jours d'incubation. Apres un sejour d'une demi-heure dans une solution de
Earle depourvue de calcium et de magnesium, le blastoderme a ete debarrasse
de son endoderme, puis de son ectoderme. Les divers greffons ont ete decoupes,
puis conserves dans du liquide de Tyrode jusqu'au moment de l'implantation.
Les somites ont ete preleves sur une longueur de 4 a 5 somites sur des embryons
de Poulet ayant recu prealablement une injection de thymidine tritiee ou sur
des embryons de Caille.
Le mesoderme somatopleural a ete preleve soit dans la zone proximale des

Developpement du plumage dorsal. I 317
lames laterales, jouxtant les somites, soit dans la zone distale des lames laterales,
a une distance du bord externe des somites approximativement egale a une lar-
geur de somite (Fig.
1
A, B).
Ann de reconnaitre l'orientation du greffon au cours de la transplantation,
une marque de noir animal a ete deposee a l'extremite anterieure dorsale de
chaque greffon.
(c) Marquage du mesoderme somitique a la thymidine tritiee
Huit heures avant le prelevement des greffons somitiques, une goutte de
liquide de Tyrode contenant 1,2 /^Ci de thymidine tritiee a ete deposee sur le
blastoderme.
{d) Marquage
biologique
Les cellules embryonnaires de la Caille (Coturnix coturnix japonica) se
distinguent des cellules embryonnaires de Poulet par la presence de 1 a 3 gra-
nules d'ADN nucleolaire fortement Feulgen-positifs (Le Douarin
&
Barq, 1969).
Ce caractere nucleaire a permis de les suivre aisement au cours des etapes de
leur deplacement dans un hote Poulet.
(2)
Techniques
histologiques
(a) Histologie consecutive aux excisions et au remplacement du mesoderme
somitique
Les embryons ont ete fixes au liquide de Bouin-Hollande 3, 24, 48, 72, 96 h
apres l'intervention. Les coupes, d'une epaisseur de
5
jum,
ont ete colorees a
l'hemalun-eosine.
(b) Histologie consecutive aux greffes de somites de Caille
Les embryons ont ete fixes au liquide de Helly
3,
24,48, 72, 96 h apres l'opera-
tion. Les coupes, d'une epaisseur de
5
/tm, ont ete colorees par la reaction nu-
cleate de Feulgen et Rossenbeck.
(c) Autoradiographie
Les embryons ont ete fixes a l'alcool acetique 0, 24, 48 et 96 h apres la greffe
de somites marques a la thymidine tritiee. Le traitement autoradiographique
a ete fait selon la technique de Caro & van Tubergen (1962). Les coupes, epais-
ses de 5/«n, ont ete couvertes de l'emulsion photographique (Ilford, L4).
L'exposition a dure de 5 semaines a 6 mois. Les coupes ont ete colorees a
Fhematoxyline de Groat/eosine.
(3) Observation des effets
morphologiques
Les embryons non soumis a
1'etude
histologique ont ete fixes 8 jours apres
l'operation, soit vers 10,5 jours d'incubation, au liquide de Bouin-Hollande.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
1
/
30
100%