Vascularisation artérielle du thalamus humain: groupes artériels extra

1
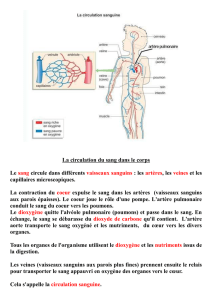
Vascularisation artérielle du thalamus
humain: groupes artériels extra-
parenchymateux
A. COSSON, L. TATU, F. VUILLIER, B. PARRATTE, M. DIOP ET G. MONNIER
Introduction
Les données récentes de l’imagerie ont permis d’importants progrès diagnostiques
dans le domaine de la neurologie vasculaire. Force est de constater cependant que
certaines interprétations restent délicates parce qu’elles reposent sur des données
morphologiques qui n’ont pas suivi la même cinétique. Ainsi en est-il de la
vascularisation du thalamus dont la pathologie confronte fréquemment le clinicien
et le neuro-radiologue à des problèmes décisionnels induits par le caractère
aléatoire de nos connaissances anatomiques.
La vascularisation du thalamus est un problème débattu depuis de nombreuses
années. Les références anatomiques concernant la constitution de l’ensemble des
groupes artériels impliqués dans la vascularisation thalamique sont
contradictoires. Elles montrent une diversité importante et des incertitudes dans la
description des apports artériels et la complexité de ces descriptions interdit toute
application concrète en pathologie.
Ces constatations nous ont incité à entreprendre, dans un premier temps, une étude
de la disposition extra-parenchymateuse des groupes artériels thalamiques, dans
un souci de clarification terminologique et de standardisation. Elle représente une
étape obligatoire avant d’aborder l’étude de la vascularisation intra-
parenchymateuse des groupes nucléaires thalamiques.
Matériel et méthodes
Ce travail a été réalisé à partir de 12 encéphales humains adultes prélevés
rapidement après le décès, soit au total 24 hémisphères (8 femmes, 4 hommes, 52-
96 ans). Les cadavres utilisés dans cette étude provenaient de dons du corps

2
enregistrés au Laboratoire d’Anatomie de la faculté de Médecine de Besançon
entre septembre 2001 et juin 2002.
Neuf encéphales ont été injectés avec un mélange de latex néoprène et d'encre de
Chine rouge et 3 encéphales avec un mélange de gélose et d'encre de Chine noire.
Les injections intra-vasculaires au latex néoprène ont été effectuées après
prélèvement de la tête entière sur des cadavres embaumés par une solution de
liquide de Winckler. Les artères carotides internes et vertébrales ont été
systématiquement injectées. La polymérisation du produit s'effectuant à froid, les
pièces anatomiques étaient ensuite plongées dans une solution de formol à 15 %,
et placées au réfrigérateur à une température comprise entre 1 et 5°C. L’encéphale
était prélevé ultérieurement. Pour l’injection à l’encre de Chine gélosée, les
encéphales étaient prélevés sur des cadavres non embaumés, dans les 12 heures
suivant le décès. Après l’injection intra-vasculaire des artères carotides internes et
vertébrales, la fixation de l'encéphale s'effectuait dans une solution de formol à
10% pendant une période de 30 à 45 jours.
Le travail de dissection a été effectué à l'aide d'un microscope opératoire et à
l’aide d’instruments adaptés à la microchirurgie. Les photographies ont été prises
avec un appareil photographique numérique branché en dérivation sur le
microscope. Les 12 encéphales ont été disséqués selon un protocole reproductible.
Les constituants du polygone artériel de la base du cerveau ont été individualisés.
Les différents troncs artériels participant à la vascularisation du thalamus ont
ensuite été repérés depuis leur origine. Les artères cérébrales postérieures (ACP),
communicantes postérieures (AComP), choroïdiennes antérieures (AChA),
choroïdiennes postéro-médiales (AChPM) et choroïdiennes postéro-latérales
(AChPL) ont ainsi été systématiquement disséquées de proche en proche. Il a
ensuite été nécessaire de sectionner le tronc cérébral à hauteur du pont et de
procéder à l’ablation du cervelet. Les lobes temporaux ont progressivement été
disséqués pour dégager la substance perforée antérieure et la fissure choroïdienne.
La corne temporale du ventricule latéral était finalement ouverte. La région des
corps géniculés et du pulvinar était alors disséquée. La paroi dorsale du troisième
ventricule était finalement abordée par dissection du mésencéphale pour visualiser
les points d’entrée des artères choroïdiennes postéro-médiales. Une section

3
horizontale de la partie supérieure des hémisphères cérébraux permettait d’aborder
les ventricules latéraux. L’ablation progressive du corps calleux puis du fornix
faisait apparaître les branches des artères choroïdiennes et permettait de suivre
leur trajet sur la face supérieure du thalamus.
Résultats
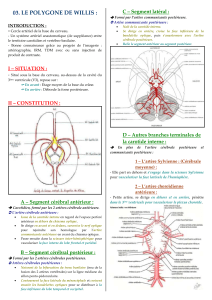
Terminologie de la description des groupes artériels
Nous avons utilisé la division en 4 segments de l'ACP, proposée initialement par
Krayenbühl et Yasargil [16] et précisée par Zeal et Rhoton [40]. Le segment P1
était compris entre l'origine de l'ACP et la jonction avec l'AComP. Le segment P2
correspondant au trajet de l'artère autour du pédoncule cérébral jusqu'au pulvinar,
était divisé en deux parties égales, une moitié antérieure appelée P2A et une
moitié postérieure appelée P2P. Le segment P3 s'étendait du pulvinar au sillon
calcarin, point habituel de division de l'artère en ses deux branches terminales. Le
segment P4 était défini par l'épanouissement de l'ACP en ses branches corticales.
L'artère pré-mamillaire a été définie comme la plus volumineuse des branches
perforantes de l'AComP [24, 35]. Elle pénétrait la substance perforée latéro-
mamillaire (SPLM), région située à l’angle du tractus optique et du pédoncule
cérébral en regard du corps mamillaire.
Les artères thalamo-perforantes étaient identifiées comme les artères issues du
segment P1 de l'ACP ou de la bifurcation de l'artère basilaire et constituaient le
groupe supérieur des artères interpédonculaires [8]. Elles pénétraient la substance
perforée postérieure rostrale (SPPR).
L'AChPL était définie comme un tronc artériel issu de l'ACP, se divisant en une
branche médiale et une branche latérale [34]. La branche latérale entrait dans la
corne temporale du ventricule latéral (VL) et suivait les plexus choroïdes en
s'anastomosant avec les branches plexuelles de l'AChA. Elle donnait des
perforantes pour la face supérieure du thalamus. La branche médiale avait un
trajet plus court, contournant le pulvinar pour rejoindre les plexus choroïdes du
VL. Elle pouvait également fournir des rameaux thalamiques.

4
L’AChPM était définie comme un tronc artériel issu de l'ACP se dirigeant vers la
partie postérieure du toit du troisième ventricule (V3). Nous avons utilisé la
division en deux branches proposée par Plets et Van den Bergh [31] : une branche
médiale pour les plexus choroïdes et une branche latérale donnant
préférentiellement des rameaux pénétrant la région péri-habénulaire. La branche
médiale s'anastomosait avec l'AChPL au niveau du foramen interventriculaire.
Le terme d'artères thalamo-géniculées a été appliqué aux artères entrant dans la
région péri-géniculée située au pourtour et entre les corps géniculés. Elles étaient
issues des artères passant à proximité de cette zone : ACP, AChPM, AChPL ou
artère colliculaire [8, 35, 40].
Le trajet de l’artère choroïdienne antérieure (AChA) a été divisé en un segment
cisternal et un segment plexuel après son entrée dans la corne temporale du VL
[35]. En accord avec Plets et al. [30,31] et Schlesinger [34], nous avons considéré
que les branches de l'AChA traversant le tractus optique ou la partie inféro-
latérale de la substance perforée antérieure (SPA), participaient à la
vascularisation du globus pallidus et de la capsule interne, mais pouvaient
également vasculariser la partie latérale du thalamus. Nous avons par ailleurs
admis que les branches plexuelles de l'AChA pouvaient émettre, dans la corne
temporale du VL, des perforantes pour le corps géniculé latéral ou le thalamus, en
particulier le pulvinar [18, 21].
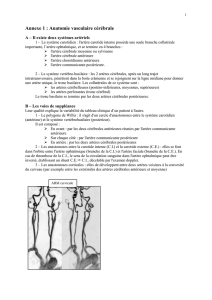
Les points d’entrée des pédicules thalamiques dans le diencéphale sont précisés
sur les figs. 1 et 2.
Artère pré-mamillaire
Il existait une artère pré-mamillaire par hémisphère, sauf pour un hémisphère, où
il en existait 2, soit un total de 25 artères : 13 naissaient du tiers moyen de
l'AComP, 6 du tiers antérieur de l'AComP, 2 de l'ACP (P1 ou P2A), 3 du tiers
postérieur de l'AComP et 1 se terminait en AComP (fig. 3).

5
Artères thalamo-perforantes
Toutes les artères thalamo-perforantes naissaient du segment P1 de l'ACP ou, en
cas de disposition fœtale de l'AComP, du reliquat artériel entre l'artère basilaire et
l'AComP. Pour 8 encéphales, la naissance des artères thalamo-perforantes était
unilatérale (fig. 4). Il existait 21 artères thalamo-perforantes au total, se divisant
en 45 branches pénétrant la SPPR, soit une moyenne de 3,75 branches artérielles
par cerveau (extrêmes 2-5).
Rameaux perforants de l'artère choroïdienne postéro-
médiale
Il existait une AChPM par hémisphère, sauf pour 2 hémisphères où il en existait 2,
soit un total de 26 artères : 17 AChPM naissaient du segment P2A de l’ACP, 4 du
segment P3, 3 du segment P2P, 2 de la jonction des segments P2P et P3. Les 26
AChPM donnaient 79 rameaux perforants pour le thalamus : 69 pour la face
médiale, 7 pour le pulvinar, 2 pour la face supérieure et 1 pour le pôle antérieur.
Le nombre moyen de rameaux par hémisphère était de 3,3 (extrêmes 0-8) (fig. 5).
Rameaux perforants de l'artère choroïdienne postéro-
latérale
Il existait 39 AChPL soit une moyenne de 1,6 artères par hémisphère (extrêmes 0-
3). Les 39 AChPL naissaient de l'ACP : 19 de la jonction des segments P2A et
P2P, 8 d'une branche corticale, 5 du segment P3, 4 du segment P2P, 3 de la
jonction des segments P2P et P3. Ces AChPL donnaient 104 rameaux pour le
thalamus : 89 pour la face supérieure et 15 pour le pulvinar soit une moyenne de
4,3 par hémisphère (0-10).
Artères thalamo-géniculées
Il existait 269 artères thalamo-géniculées, soit une moyenne de 11,2 par
hémisphère (extrêmes 7-14) : 143 artères naissaient de l'ACP ou d'une de ses
branches corticales, soit une moyenne de 5,9 artères par hémisphère, 64 d'une
AChPM, 53 d'une AChPL et 9 d'une artère colliculaire (Fig. 6).
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%