
Accueil
B.T.S. Année 1
B.T.S. Année 2
Licence Pro
Master Pro
Liens
Livres
Foire Aux Questions F.A.Q.
Curriculum Vitae
Contactez-moi
Microbiologie
Multiplication des bactéries et des champignons
Etude de la croissance
Introduction
Chez les micro-organismes unicellulaires, la croissance correspond à la multiplication des
individus. L'étude de la croissance revient donc à évaluer l'augmentation de la population et
s'effectue en analysant la courbe de croissance d'une culture microbienne.
Après avoir étudié les différentes techniques d'étude de la croissance:
- évaluation de la biomasse (poids sec, turbidimétrie)
- évaluation du nombre de cellules (dénombrement en lame de Malassez, mise en culture et
comptage des colonies)
- évaluation de l'activité cellulaire (consommation d'un substrat, production d'un produit).
Nous étudierons une courbe de croissance en milieu non renouvelé (en «batch»), et nous
présenterons les cultures en batch alimenté et en continu.
1. Méthodes d'études de la croissance
Mesure de la biomasse
Dans une population bactérienne en croissance, la biomasse (ou quantité de matière
vivante) augmente régulièrement. On peut l'évaluer de différentes façons.
Détermination du poids sec

II s'exprime en g/L de culture. C'est la méthode de référence, car la plus précise quand elle
est bien maîtrisée, mais elle est délicate à mettre en oeuvre. (masse de microorganismes
complètement déshydratés dans une culture).
Mesure d'une absorbance
Opacimétrie
Le trouble d'un milieu ou sa densité optique est directement proportionnel à la
concentration cellulaire dans le milieu, selon la loi de Beer-Lambert:
A = Ɛ x c x l
Les mesures d'opacimétrie se font entre 550 et 650 nm.
Avantages: Simples à mettre en oeuvre; Peu couteux.
Inconvénients:Domaine de linéarité : λ= 650nm; A linéaire jusqu'à A = 0,5 --> A réelle du
milieu. Inapplicable lorsque le milieu est trouble.
Valeur estimée par excès: on compte ici les cellules vivantes et les cellules mortes donc on
surestime la quantité de biomasse.
Turbidimétrie:
La turbidité d'un milieu contenant des particules provoque des altérations du trajet optique
dues aux phénomènes de diffusion / réfraction.
La turbidité se mesure par détermination de l'épaisseur minimale de suspension
empêchant la vision nette d'une mire, ou par comparaison avec un étalon.
Néphélométrie:
Mesure de la diffusion de la lumière par les particules.
Mesure du nombre de cellules
Le nombre de cellules d'un milieu peut-être évalué par des méthodes directes cad compter
directement le nombre de cellules.
Comptage à l'hématimètre
Compteur de particules
C'est un système automatique qui dénombre les bactéries qui passent entre 2 électrodes au
niveau d'une micropipette.
Inconvénients: Ce système est peu adapté à la taille des bactéries. De plus il dénombre les
cellules vivantes et mortes. Donc par excès.
Des marqueurs fluorescents à l'acridine (qui se fixe à l'ADN) permettent de différencier les
cellules vivantes (vertes) des cellules mortes (rouges).

On peut également évaluer le nombre de bactéries par des méthodes indirectes cad par
culture de cellules.
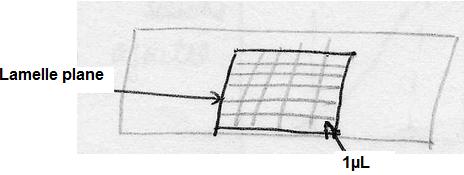
Numérations en milieu solide
Cette technique consiste à réaliser des dilutions décimales du produit à étudier et à
ensemencer soit 1 mL en profondeur d'un milieu solide en surfusion, soit 0,1 mL en surface
à l'aide d'un râteau. (2 boîtes par dilution)
Les résultats sont donnés en UFC.
Avantages des méthodes indirectes: On ne compte que des cellules vivantes.
Inconvénients des méthodes indirectes: On ne fait pas de différences entre une colonie
issue d'un microorganisme ou issue d'un amas de microorganismes. On augmente le
pourcentage d'erreurs. Il y a une étape d'incubation donc les résultats ne sont pas
immédiats.
Détermination de l'activité
II est possible de suivre par des techniques biochimiques la consommation d'un substrat (ex
: suivi de la consommation de glucose par dosage enzymatique) ou l'apparition d'un produit
(ex:dosage de l'éthanol).
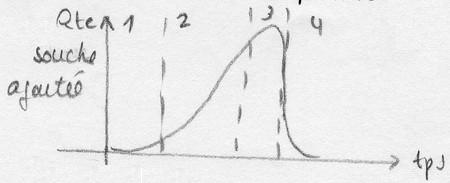
On peut également suivre l'activité des micro-organismes en étudiant le maintient d'un
paramètre régulé.
Ex: le pH est en général un paramètre régulé au cours d'une fermentation. Donc sa valeur
ne donne pas d'information sur l'activité des micro-organismes, mais l'action nécessaire
pour maintenir le pH constant sera , elle, représentative du niveau d'évolution de la
réaction de fermentation. Ainsi, en suivant la quantité de base injectée par unité de temps,
la courbe obtenue sera:
2. Suivi de la croissance en milieu non renouvelé
Un milieu non renouvelé est un milieu pour lequel on n'apporte pas de nouveau substrat en
cours de culture ainsi la croissance est limitée et elle s'arrête par épuisement des substrats

et accumulation des déchets. En pratique, c'est ce qui se produit dans un erlen ou un tube
de bouillon.
La courbe de croissance
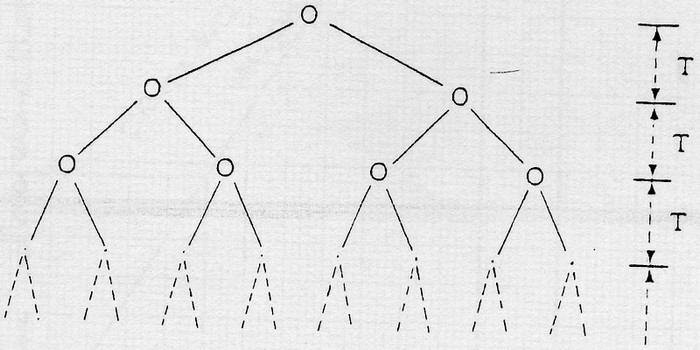
La croissance cellulaire se fait par divisions binaires successives selon le schéma:
T étant le temps s'écoulant entre 2 divisions.
On peut écrire l'équation suivante:
N: nombre de cellules totales
N0: nombre de cellules totales à t=0
N = N0 x 2t/T
=> ln N = ln N0 + t/T ln 2
ln N = ln N0 + ln 2 / T x t
avec ln N0 = constante et ln 2 / T = variable car T est fonction des conditions de cultures.
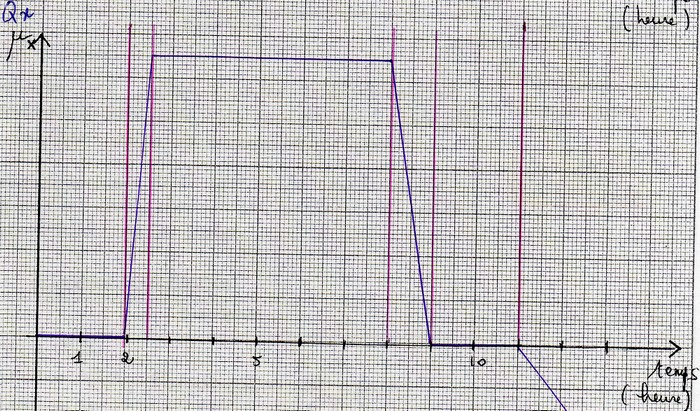
Les différentes phases de la croissance

 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}