Cours de Génomique Structurale : Chromatines et Expression Génique
Telechargé par
IMAD AGNAOU

Pr. Izaabel Hassan e mail : h.izaabel@gmail.com
Cours de Génomique structurale
•La génomique est l'étude exhaustive du génome, des différentes
séquences qui le compose et en particulier, de l'ensemble des
gènes, de leur structure, de leur disposition sur les chromosomes,
de leur séquence, de leur fonction et de leur rôle.
• Références :
• Génétique (8eédition) de W. Klug, M. Cumming et C. Spencer
• Gènes V de Lewin
• Précis de génomique de Gibson & Muse
• Expression du gènome de Herbomel
• Gènes & Gènome de Singer et Berg
• Biologie moléculaire et médecine de J.C Kaplan et M. Delpech
• Génétique moléculaire Humaine de T. Strachan et A.P. Read
• Introduction à l’analyse génétique de Griffiths, Miller et al
Chapitre I
Structure fine de la chromatine
et régulation de l’expression
génique.

•Chapitre I : Structure fine de la chromatine et
régulation de l’expression génique.
I. I. Généralités
Dans le noyau :
Dans le noyau :
–
–le DNA n
le DNA n’
’est jamais nu,
est jamais nu, mais toujours
mais toujours
associ
associé
éà
àdes prot
des proté
éines et même
ines et même à
àdes
des
ARN.
ARN.
–
–L
L’
’ensemble constituant ce que l
ensemble constituant ce que l’
’on
on
appelle
appelle la chromatine
la chromatine.
.
La vie cellulaire impose qu
La vie cellulaire impose qu’à
’à tout moment
tout moment
l
l’
’information ad
information adé
équate soit trouv
quate soit trouvé
ée parmi
e parmi
les 3 milliards de paires de bases, puis
les 3 milliards de paires de bases, puis
lue de mani
lue de maniè
ère r
re ré
égul
gulé
ée.
e.

•
•Cette information doit être
Cette information doit être prot
proté
ég
gé
ée
e,
,
r
ré
épar
paré
ée
een cas d
en cas d’
’alt
alté
ération et enfin
ration et enfin
p
pé
érennis
rennisé
ée
edans les cellules filles.
dans les cellules filles.
•
•Tout cela est l
Tout cela est l’œ
’œuvre
uvre -
-au moins en tr
au moins en trè
ès
s
large part
large part -
-des prot
des proté
éines de la
ines de la
chromatine
chromatine qui joue un rôle centrale.
qui joue un rôle centrale.
•
•Ce n
Ce n’
’est que par la connaissance
est que par la connaissance
d
dé
étaill
taillé
ée de leur structure que l
e de leur structure que l’
’on
on
pourra comprendre
pourra comprendre la r
la ré
égulation de
gulation de
l
l’
’expression des g
expression des gè
ènes.
nes.
La chromatine
La chromatine se pr
se pré
ésente
sente (au stade
(au stade
interphasique)
interphasique) le plus souvent sous la forme
le plus souvent sous la forme
d'une mati
d'une matiè
ère sans structure particuli
re sans structure particuliè
ère.
re.

A certains moments de la vie de la cellule
A certains moments de la vie de la cellule (aux
(aux
moments des multiplications (ou stade
moments des multiplications (ou stade
m
mé
étaphasique),
taphasique), la chromatine perd son aspect
la chromatine perd son aspect
diffus et se condense en structures bien
diffus et se condense en structures bien
d
dé
éfinies:
finies: les chromosomes.
les chromosomes.
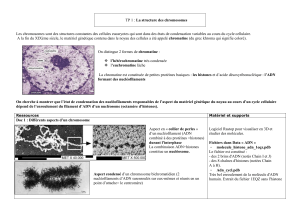
La fibre de la chromatine observée au microscope électronique a un
diamètre de 100nm.
Dans certains conditions expérimentales cette fibre peut présenter soit
un diamètre de 30nm (300A°) ou 10nm (100A°).
Ces différentes valeurs sont très supérieures au diamètre de la double
hélice de l’ADN qui est de 20A°.
100 nm 30 nm 10 nm

Chapitre I : Structure fine de la chromatine et
régulation de l’expression génique.
I. I. Généralités
I. II. Structure de base de la fibre chromatinienne.
1. Structure du Nucléosome
L’ADN est coupé en
des fragments dont
la taille est un
multiple d’une unité
de longueur, après
fractionnement par
électrophorèse sur
gel d’agarose, on
obtient une échelle
s’étendant sur une
dizaine de bandes
successives de
l’ordre de 200 pb.
Digestion de la chromatine par la Nucléase de Microcoque.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%