3 éme année licence Biologie et physiologie végétale Biotechnologies Végétale
1
SALHI N.
Partie I : Bases de la multiplication
I-LES MÉRISTÈMES PRIMAIRES
1- Définition
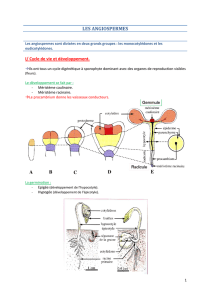
L’embryon d’une Angiosperme comporte déjà les ébauches des futurs méristèmes (du grec meris =
partie et stêma = filament) caulinaires (des tiges) et racinaires.
Ces méristèmes primaires, qui sont à l’origine des tissus de la plante, sont situés chez les
Spermaphytes, au niveau de la gemmule et de la radicule. (fig.1 a et b)
De la germination jusqu’à la mort de la plante les méristèmes assurent la croissance en longueur des
tiges et des racines.
Même si cette croissance se ralentit parfois, il est possible de parler de croissance indéfinie. Ces
méristèmes primaires en fonctionnant vont donner des tissus. Ils sont dénommés tissus primaires pour les
différencier des tissus secondaires qui apparaissent chez certaines plantes ultérieurement
Fig 1 : a- coupe longitudinale dans une graine b- Emplacement des méristèmes I
2- La cytologie des cellules méristématique :
Les cellules du méristème primaire sont petites et isodiamétriques. Elles sont parfaitement
jointives (pas de méats). Elles possèdent un noyau central occupant une partie importante du
volume cellulaire. L’appareil vacuolaire est réduit et il est constitué par de très petites vacuoles qui
sont soit sphériques soit disposés en un très fin réseau. Les mitochondries sont nombreuses et il
n’existe pas de plastes différenciés. ( tab 1)

3 éme année licence Biologie et physiologie végétale Biotechnologies Végétale
2
SALHI N.
Tableau 1 : Caractéristiques des méristèmes primaires.
3- Fonctionnement des méristèmes
Avant de commencer quelques précisions terminologiques :
• L’apex est une notion morphologique qui correspond à l’extrémité d’une tige ou d’une racine.
• Le méristème est le tissu indifférencié qui assure la production d’organes nouveaux (feuilles, racines,
fleurs).
• Un méristème apical se trouve à l’extrémité d’une tige ou d’une racine donc à l’apex. Aux apex
apparaissent les nouveaux organes grâce au fonctionnement des méristèmes. Ceux-ci sont de deux types : les
méristèmes apicaux caulinaires à l’origine des parties aériennes et les méristèmes apicaux racinaires pour
l’appareil souterrain. Ils fonctionnent de manière très différente.
a- Le fonctionnement du méristème caulinaire
Le méristème caulinaire (de la tige) est responsable de l’édification de la partie aérienne de la plante. De
lui, apparaissent des cellules qui en se multipliant et en se différenciant donneront les feuilles, les bourgeons
axillaires et floraux et les tiges. De manière, tout à fait répétitive et indéfinie, jusqu’à la mort de la plante, il
engendre la formation des phytomères, constitués d’un axe (ou entre-noeud) d’une ou plusieurs feuilles (cas
des feuilles opposées comme chez l’érable) et d’un bourgeon axillaire.
Chez les Angiospermes, ce méristème forme un dôme (plus rarement une structure plate), de 0,5 à 3 mm
de diamètre, composé de cellules de petite taille non équivalente.
la section centrale du méristème révèle, après coloration des ARN au vert de méthyle-pyroxile (qui permet
de montrer l’activité métabolique des cellules), trois régions (fig. 2) :

3 éme année licence Biologie et physiologie végétale Biotechnologies Végétale
3
SALHI N.
Fig 2 : Différentes zone du méristème caulinaire
• Une zone axiale(Za), très peu active quant aux mitoses, avec deux couches superficielles, les
tunicas T1 tout autour du méristème se distingue par ses divisions strictement anticlines (cloisons
perpendiculaires à la surface). Elle est à l’origine de l’épiderme. La tunica T2 initie les feuilles par
cloisonnements périclines.et le corpus, C, Située au-dessous donne les tissus centraux de la tige et
des feuilles. Dans cette couche, les mitoses s’effectuent dans tous les sens.
• Une zone latérale(ZL), entourant cette zone axiale, Za, la partie à droite correspond à l’apparition
d’une feuille(ZLF), On distingue des divisions périclines, dp, (les cloisons sont parallèles à la
surface)
• Un méristème médullaire, (Mm), aux mitoses peu fréquentes formant des files empilées de
cellules à l’origine de la moelle centrale, M.
Nb : (Dans la littérature anglo-saxonne, T1 et T2 sont remplacés par L1 et L2, L étant
l’abréviation du mot anglais layer (couche). )

3 éme année licence Biologie et physiologie végétale Biotechnologies Végétale
4
SALHI N.
La formation d’un phytomère comprend l’apparition d’une ou plusieurs feuilles, du ou des bourgeons
axillaires et de l’allongement de la tige. Chaque étape correspond à un fonctionnement particulier de l’apex
caulinaire :
• L’initiation foliaire correspond à un élargissement de la zone latérale ZLF qui produit un initium foliaire.
Provenant de divisions essentiellement périclines, cet initium à une forme de croissant qui par surrection va
grandir et donner l’ébauche foliaire (fig. 3). Le fonctionnement du méristème est donc périodique, il se
produit une alternance régulière entre une croissance horizontale (initium foliaire) et verticale (ébauche
foliaire).
• Le bourgeon axillaire est d’origine superficielle exogène, car c’est dans les cellules des tunicas qu’il se
différencie.
• La formation de la tige est liée à l’activité du méristème médullaire, au centre du dôme.
b- La transformation du méristème apical en méristème floral
Le stade de développement physiologique de la plante peut entraîner des modifications d’activité du
méristème caulinaire. Le changement le plus spectaculaire correspond à la floraison.

3 éme année licence Biologie et physiologie végétale Biotechnologies Végétale
5
SALHI N.
Sous l’action de signaux encore mal identifiés, mais qui pourraient être liés à deux protéines
sensibles à la lumière, un phytochrome et un cryptochrome, le méristème caulinaire se transforme
soit en méristème floral, à l’origine d’une fleur unique, soit en méristème d’inflorescence, qui à son
tour produira des méristèmes floraux.
Ce changement s’accompagne de modifications exceptionnelles : un système à croissance indéfini
comme le méristème apical, devient défini, puisque, une fois la morphogenèse florale terminée, le
méristème arrête toute activité et disparaît à la chute des fleurs ou des fruits.
Ces transformations correspondent à un ralentissement d’activité de la zone latérale (ZL) qui
pourtant donnera les sépales, premières pièces florales apparaissant, tandis que le corpus prolifère
abondamment en mettant en place ce qui deviendra le réceptacle floral.
La tunica 2 sera à l’origine des pièces florales reproductrices d’ou son nom de proméristème
sporogène.
c- fonctionnement du méristème racinaire
L’allongement de la racine se fait par son extrémité au niveau du méristème racinaire. Celui-ci est d’un
fonctionnement plus simple que le méristème caulinaire. À l’extrémité des racines, on distingue une coiffe,
qui protège le méristème .Entre celle-ci et les poils absorbants, on observe une zone quiescente (sans division
cellulaire), autour de laquelle on peut remarquer de très nombreuses divisions cellulaires qui sont à l’origine
des tissus de la racine (fig4.).
Fig 4 : Méristème apical racinaire (a) extrémité d’une racine, (b) coupe longitudinale de
l’extrémité d’une racine observée au microscope optique (c) le méristème apical racinaire
avec les principales zones.
 6
7
6
7
1
/
7
100%