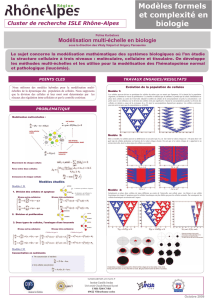
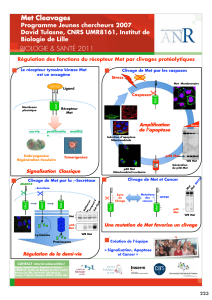
Culture Cellulaire Végétale In Vitro : Dynamique et Régulation
Telechargé par
zahradine67

Dynamique et régulation cellulaire :
La culture cellulaire in vitro permet de créer, à partir d'un petit bout de plante, un
grand nombre de plantes du même
type.
Par exemple, sur une pomme de
terre qui bourgeonne à l'obscurité,
on peut récupérer les pousses, le
fragmenter, le mettre en culture et
récupérer un milliers d'individu
provenant d'une même pousse.
Les méthodes de culture cellulaire
sont multiples et dépendent des
espèces. On peut utiliser un
embryon de plantes, un organe, un
cal (cellule indifférencié) ou une
cellule par exemple.
Cultures cellulaires végétales : Historique et impact :
1. Totipotence des cellules végétales :
L'idée que le noyau d'une cellule était le site pouvant régénérer tout un organisme à
été émis par Haberlandt en 1902 : « Toute cellule végétale est capable de régénérer
un nouvel organisme identique à celui dont elle est issue.» (pas vraiment possible sur
toutes les cellules)
De nombreuses cellules végétales sont totipotentes tandis que seul le zygote est
totipotent chez les métazoaires.
En 1965, Vasil et Hildebrandt on pu prouver que l'idée de Haberlandt était juste en
régénérant un plant de tabac.
2. Historique de la culture in vitro :
En 1934, White réussit à créer une culture de tomates en milieu in vitro via une racine
de tomate. (Le milieu était composés d'eau, de sels minéraux, d'extrait de levure, du sucre et de
l'auxine, seul hormone végétale connue à l'époque.)

En 1939, Gautheret a permis le développement indéfini (On cultive aujourd'hui encore
des cellules du premier cal de Gautheret) d'un cal de cellules dédifférenciées à partir
d'un fragment de racine de carotte. Il s'agit de la première vrai culture in vitro
(Dans du verre). Les cellules ayant poussé à partir de la racine différencié sont des
cellules indifférenciés.
En 1962, Murashige et Skoog mettent au point le premier milieu de base pour la
culture in vitro :
- Sels minéraux (macro et micro-éléments en g/L et μg/L)
- Sucres (saccharose)
- Vitamines (B)
- Hormones (Auxine et cytokinines)
- pH acide à 5,5
La culture est faite à température, lumière et hygrométrie contrôlée.
Via cette technique, on a pu faire de la prolifération de méristème de tige, ce qui
n'avait jamais été fait auparavant, le méristème ayant toujours été réfractaire à la
multiplication végétative in vitro.
Des substances organiques sont ajoutés parfois pour tenter de booster la croissance de plante, les
espèces végétales ne répondant pas tous de la même façon à la culture in vitro (acides aminés, extrait
de banane, pomme de terre, etc. )
3. Activation de processus cellulaire programmés :
Le fragment sectionné, blessé, doit cicatrisé pour ensuite régénérer spécifiquement
le morceau endommagé.
•1ère étape : Dédifférenciation cellulaire : Une fois un fragment coupé et retiré
de l'organisme mère, il perd les signaux de position, nutrition, dominance apical
de l'auxine, etc. qu'il avait auparavant. Il redevient une cellule dédifférencié
de cals.
•2ème étape : Multiplication cellulaire
•3ème étape : Redifférenciation cellulaires

Certains résultats sont meilleurs :
➢Quand les tissus d'origine du tissus de départ sont des tissus jeunes
(embryons zygotiques immatures, hypocotyles (tissus en dessous du cotylédon)
et cotylédons (pseudo-feuille poussant au début du développement de la
plante) de germinations, méristèmes de plantes adultes).
➢Quand les cellules sont diploïdes plutôt que polyploïdes.
➢Chez certaines familles de plantes (Solanacées, Brassicacées (4 pétales
formant une fleur (A. Thaliana, Choux, Carotte))) et certains génotypes au sein
d'une famille (Tournesol)
4. Développement d'explants pluricellulaire :
•1ère voie : L’organogenèse : Passer d'un cal (cellules indifférenciés) à une
plantule. En partant d'embryons immatures, on peut générer des pousses de
tiges ou des racines selon le milieu de culture dans lesquelles on les placent.
•2ème voie : Embryogenèse somatique : On part d'un explant immatures pour
donner un 'organe' ayant une morphologie d'embryons végétales, appelés
embryons somatiques. Ces embryons permettent ensuite de donner des
individus. C'est le développement initial d'un tissu indifférencié de type
embryonnaire qui se développe en un individu complet.

5. Conditions de régénération de plantules :
Le milieu de culture se doit d'être :
1. Aseptisation du milieu ET de l'explant.
2. Milieu de culture solide (agarose)
•AVANTAGES : Manipulation facile, action physique du maillage favorise les divisions et l'ancrage.
•LIMITES : Diffusion des nutriments
Milieu de culture liquide
•AVANTAGES : Absorption aisée des nutriments
•LIMITES : Manque d'oxygénation (besoin d'agitation), sécrétion de produits toxiques dans le milieu
Les repiquages réguliers sont indispensables.
Les risques sont la perte de stérilité et la contamination par des bactéries ou levure.
Rôle des hormones dans la croissance :
L'auxine, de son vrai nom Acide indole-3-acétique (AIA) est synthétisé dans l'apex, le
méristème apical des pousses. Ces effets sont les suivants :
- élongation de la tige
- différenciation et croissance des racines (en grande quantité)
- maturation du fruit
- inhibition de croissance des bougeons latéraux
- phototropisme des feuilles et géotropisme des racines
La cytokinines, a d'abord été appelé zéatine puis fut synthétisé en laboratoire. Elle
est synthétisé dans les racines et les tissus à croissance rapide. Ses effets sont les
suivants :
- retard de la sénescence
- ramification et perte de domination apical (antagoniste de l'auxine)
- dépendant du taux d'auxine associé
Choix de la balance
hormonale :
Quand on a une
majorité d'auxine, la
croissance des racines
est privilégié. Quand on
a une majorité de
cytokinines, on a un
développement des
bourgeons végétatifs.
Enfin, si on a une
quantité faible d'auxine
et de cytokinines, on
obtient une floraison.

6. Applications de la culture in vitro :
Les applications de la
culture in vitro sont
nombreuses, on peut faire
du 'sauvetage
d'embryons', de
l'haplodiploïdisation,
l'obtention de
protoplastes, la
multiplication conforme ou
la culture de méristème.
Culture de méristèmes :
En 1952, Morel et Martin développe une technique pour protéger et sauver les plantes
malades, touchés par des virus.
On cultive le méristème car c'est la seule zone qui n'est jamais contaminé par un
virus. Il pousse plus vite que le virus n'infecte les cellules.
Sauvetage d'embryons :
Certains croisement interspécifiques ne permettent pas le développement de graines !
Un embryon crée en forçant un croisement entre deux plantes donne un hybride que
l'on n'aurait pas obtenu en croisant pollen et anthère.
La culture des embryons immatures permet l'obtention de plantes.
Les avantages sont que l'ont peut s'affranchir de l'étape de dormance des
semences, le développement des embryons est homogène et l'accélération du cycle
végétatif.
Haplodiploïdisation :
Mise en culture des organes reproducteurs afin de développer des individus à partir
des cellules reproductrices et gamètes. Ces cellules contiennent une seule copie de
l'information génétique.
Au cours de la régénération, on peut réussir à doubler à l'identique cette copie. Les
individus obtenus sont des lignées pures car ils ont la même information sur les deux
chromosomes, ils sont homozygotes.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
1
/
65
100%